Is there fossil evidence of 'missing links' between humans and apes? Did ancient humans live millions of years ago?
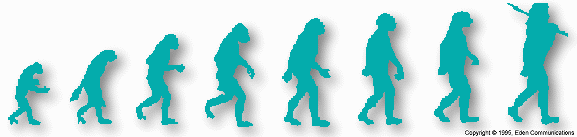
In tracing the alleged evolution of humans from primate ancestors, palaeoanthropologists have been frustrated by the existence of two extensive gaps In the fossil record. The first gap, known as the hominoid gap, goes from 32 to 22 million years ago on the evolutionary time-scale. This period is important because it is during this period that evolutionists believe that the ancestors of apes and humans were splitting off from the tine leading to the monkeys. Yet, with one possible exception, [1] this time-frame is devoid of hominoid fossil evidence to document this alleged divergence.
The second gap, known as the hominid gap, extends from 14 to 4.5 million years ago. This second period is equally critical for evolutionary theory because it is the time when the ancestors of the australopithecine and human group were allegedly diverging from the ancestors of the African apes, especially the chimpanzees. This period is also devoid of any relevant fossil material, except for a few fossil scraps too fragmentary for diagnosis. Thus, the recent news that the hominid 'missing link' has been discovered was greeted with great exuberance by the evolutionist community.
The “Missing Link”
What is claimed to be one of the most sensational fossil discoveries involving human origins was made by Tim D. White (University of California, Berkeley), Gem Suwa (University of Tokyo), and Berhane Asfaw (Ethiopian Ministry of Culture), Known first as Australopithecus ramidus [2] and later changed to Ardipithecus ramidus, [3] these fossils were found in sedimentary Pliocene deposits at Aramis, Middle Awash, northern Ethiopia between December 1992 and December 1993.
The fossil discovery is made up of associated and isolated adult teeth, a child's mandible fragment, two partial cranial bases, and seven fragments from a left arm. These 17 fragments were found in association with other primate and vertebrate fossils, Radioisotopic dating, geochemical analysis, and biochronological considerations are said to suggest a date of 4.4 million years ago. [4] (In late December 1994 a mandible and partial postcranial skeleton were found of what is thought to be the same type of individual. Details of this discovery have yet to be published. [5])

Evolutionists have been unstinting in their praise of the discovery of Australopithecus ramidus However, there is reason, to challenge their claims and to suggest that the fossils actually represent a form of pygmy chimpanzee. It is significant that, according to evolutionists, no fossils of chimpanzee have ever been found. Some have suggested that fossil chimpanzees and other primates actually have been found but they were interpreted as human ancestors rather than as fossil primates.
A contrast is seen between the report of Australopithecus ramidus in Nature and the reports of this discovery In the popular media. One senses a note of caution in the heading of the Nature article: 'The antiquity and primitive morphology of Australopithecus ramidus suggests that it represents a long-sought potential root species for the Hominidae' [6] The popular press has not reflected that caution. Colin Groves (Australian National University) said in The Canberra Times: “…the missing link is no longer missing”. [7] Time magazine writes: 'Bones from the Ethiopian desert prove that human ancestors walked the Earth 4.4 million years ago' [8] Newsweek states: ‘Ramidus confirms once and for all that the common ancestor lived just a little more than 4.4 million years ago.’ [9]
Lines of Evidence for Australopithechus Ramidus
The facts themselves do not warrant the certainty found in the popular press. Five lines of evidence are given for believing that Australopithecus ramidus is the hypothetical 'missing link'—the common ancestor of humans and chimpanzees. The first one is stated by Colin Groves. He claims that creationists cannot say that Australopithecus ramidus is just an ape because:
“It doesn't exist alone: it's in a context of all those hundreds of other pre-human remains. The ones that are slightly younger than ramidus are slightly more human like; those that are younger still are more human like still, and so on. It's a graded series from then to now.” [10]
Groves cites the various fossil categories that make up the progression from modern humans back to Australopithecus ramidus In this progression he lists the Neanderthals, Homo erectus, Homo ergaster, Homo rudolfensis, Homo habilis, A Australopithecus africanus, A Australopithecus afarensis, and finally Ardipithecus ramidus. Evolutionists often use this sequential type of 'proof' because they know how impressive it is, especially in visual form.
However, the validity of that line-up depends upon the integrity of each of the fossil individuals in that line-up and the accuracy of the dates ascribed to each one. Yet, every 'fossil watcher' knows that the individuals in that line-up change from time to time. Some are dropped; others are added. That line-up has changed considerably in the last ten years. Now it is in a state of confusion. The particular line-up that Groves suggests is not the line-up accepted by all evolutionists. This 'proof' is really no proof at all. The fossils involved and the dates ascribed to them are subjectively determined and are constantly subject to change.
The second line of evidence regarding Australopithecus ramidus is its date, 4.4 million years ago. This line of evidence is linked to the first one because Australopithecus ramidus gets its prominence by being the last one in that graded series, being older than Lucy (Australopithecus afarensis). It thus extends human ancestry farther back in time. It is said to be the earliest known hominid. ('Hominid' is an evolutionary term referring to all fossil individuals who are believed to be direct human ancestors, including the australopithecines, especially those believed to have walked erect.)
It is universally understood by evolutionists that the value of a fossil to their claims is in direct proportion to its ability to be accurately dated. This demands that the geological context of the fossil be established beyond question. Time states that the Australopithecus ramidus fossils '…were enclosed in sedimentary rock that was neatly sandwiched between layers of volcanic ash ….' [11]
Newsweek says that the fossils were '…locked in 4.4 million-year-old sediment.' [12] However, White et al write: 'All hominid specimens were surface finds….' [13] There is no indication in that report or in the dating article by Wolde Gabriel et al. [14] that the discoverers were able to determine the exact places from which these fossils had eroded. There is the assumption that they eroded in the places where they were found, but there is nothing in the reports to exclude their having been washed in from elsewhere during the heavy seasonal rains. This fact, in itself, would seem to place a degree of contingency on the dating of these fossils.
In a letter to Nature regarding the date of 4.4 million years ago, [15] John Kappelman (University of Texas, Austin) and John Fleagle (State University of New York, Sunny Brook) speak of the difficulty of both radiometric and palaeomagnetic dating in that area of Ethiopia, and demonstrate that the age of Australopithecus ramidus may have been overestimated by as much as 0.5 million years ago. They suggest that Australopithecus ramidus may not be the oldest hominid but merely a contemporary of the Australopithecus afarensis (Lucy) fossils found at four different sites in Ethiopia and Kenya.
Responding to this letter, Wolde Gabriel et al admit that the starting point of their dating process involved 'biochronological comparisons' of fauna from other places. [16] In other words, an estimate was made of what the date should be based on the evolutionary development of similar fauna found elsewhere.
This situation appears to be similar to the tortured ten year attempt to date the famous fossil skull, KNM-ER 1470, discovered by Richard Leakey in northern Kenya. (KNM stands for Kenya National Museum, where that fossil is housed; ER stands for East Rudolf, the area east of Lake Turkana where the fossil was found; and 1470 is the museum acquisition number.) In the case of 1470, after failure to achieve concordance through a number of different dating techniques on four different occasions, the final determination of the date for 1470 was based on 'biochronological comparisons', the evolution of pigs in East Africa. I have written extensively on this matter showing how the dating process can be manipulated to achieve a date acceptable to the demands of evolution. [17]
A third line of evidence that Australopithecus ramidus is the 'missing link', involves the thickness of the enamel on the fossil teeth. After referring to the 'compelling' evidence that Australopithecus ramidus is the oldest hominid species yet discovered, Bernard Wood (University of Liverpool) writes of these fossils:
They represent the remains of a species that lies so close to the divergence between the lineages leading to the African apes and modern humans that its attribution to the human line is metaphorically—and literally -by the skin of its teeth. [18]
Tim White et al. explain why:
A comparison of this ratio of enamel thickness suggests that Australopithecus ramidus may be characterized as intermediate between the chimpanzee and the Australopithecus afarensis africanus early Homo conditions. [19]
However, Peter Andrews (Natural History Museum, London) disagrees:
…all other hominids, including modern humans, have relatively thick enamel… . So the thin enamel of ramidus is more of what you'd expect from a fossil chimp. [20]
White et al. are using an improper diagnostic tool in seeking to establish the 'missing link' status of Australopithecus ramidus. Several studies have shown that because of genetic variation in enamel thickness in primates and hominids, together with environmental and nutritional factors, enamel thickness measurements should not be used in seeking to establish phylogenetic relationships. [21], [22]
Another line of evidence concerns the first deciduous molar (dm1) found in the child's mandible of the Australopithecus ramidus assemblage. White et al. write:
The dm1 has been crucially important in studies of Australopithecus since the discovery of the genus 70 years ago, and has been used frequently as a key character for sorting apes and hominids. The Aramis dm1 is morphologically far closer to that of a chimpanzee than to any, known hominid. [23]
Obviously, White et al. mean that Australopithecus ramidus is closer to the chimpanzee morphology in this particular area, and hence it is the best candidate for “missing link” status. However, his statement, together with Peter Andrew's statement above, supports our contention that Australopithecus ramidus is actually a fossil chimpanzee. Given the genetic variation in chimpanzee teeth, how does one determine that a certain tooth is 'close to a chimpanzee tooth' without being a chimpanzee tooth?
The fifth line of evidence for Australopithecus ramidus as the 'missing link' is the claim that It might have been bipedal. Here, White et al admit that the evidence is tenuous and they wisely choose not to press the issue. However, the issue is important since the definition of a hominid involves bipedality. In human evolution, the advent of bipedalism signals the beginning of the human line.
Hence, White et at are anxious to establish Australopithecus ramidus as possibly bipedal. Since no foot, leg, or hip bones were discovered, the evidence hinges on the placement of the foramen magnum, the hole at the base of the skull through which the nerves of the spinal chord pass as they go from the brain to the body. In humans, because we stand erect and are bipedal, the foramen magnum is near the center of the base of the skull. In quadrupeds, it is further to the rear of the skull.
Two small fossil fragments from the base of the skull are all that Australopithecus ramidus has to offer by way of evidence. White et al. state that these fossils '…evidence a strikingly chimpanzee-like morphology…' [24]
They go on to say that the position of the foramen magnum as deduced from these skull fragments ‘…may correlate with bipedality although this remains to be demonstrated.’ [25] Bipedality is essential if Australopithecus ramidus is to be considered a hominid and the “missing link.” However, since White et al. admit that bipedality has not been demonstrated, a refutation is hardly necessary.
In each of these cases, the attempt is made to squeeze huge amounts of evolutionary information out of the tiniest of fossil fragments. The evidence for the 'missing link' seems to be more in the imagination of the palaeoanthropologists than in the fossils themselves. The evidence that Australopithecus ramidus is a hominid and the 'missing link' is simply not impressive.
A Diagnostic Problem
Henry Gee, a senior editor of Nature, presents a problem which is both practical and philosophical and which no one else, to my knowledge, has addressed. Even if we suppose for the sake of argument that human evolution is true, when we get close to the branching point of the human line from the chimpanzee line, how can one tell if a particular fossil individual is a true hominid or just a chimpanzee with a bit of genetic variation?
Speaking as an evolutionist, Gee is honest enough to admit that there is a problem in identifying whether Australopithecus ramidus is on the human line (a hominid) or on some other line: '… with creatures as primitive as Australopithecus ramidus it will be almost impossible to tell the difference.' [26]
Using the discovery of Australopithecus ramidus as proof of evolution is a classic illustration of begging the question. Australopithecus ramidus does not prove evolution. In fact, one must assume the truth of evolution in order for Australopithecus ramidus to have any significance in human ancestry. If evolution is not true, Australopithecus ramidus becomes merely a fossilized chimpanzee—which, in, the light of the evidence, is the more logical way to interpret it.
Australopithechus Ramidus and Pygmy Chimpanzees
To illustrate the extravagant claims of evolutionists regarding Australopithecus ramidus in the light of the very limited evidence, allow me to use the pygmy chimpanzee (Pan paniscus) as an illustration. The pygmy chimpanzee, also known as the bonobo, is smaller than the common chimpanzee (Pan troglodytes), is distinct from it in several ways, and is rather rare. It is found only in Zaire, south of the Zaire River, where it inhabits the humid rain-forests “of that flat river basin. It was not ‘discovered’ until the 1930s…”
Although rare, there are dozens of pygmy chimps in zoos around the world. They can be observed at the Yerkes Primate Center, Emory University, Atlanta. Many skeletons of them are available for study. They have also been studied in the wild by a Japanese team since 1974 and by an American team since 1979. Yet, there is a robust question among scientists as to the relationship of the pygmy chimp to the common chimp. Although the two are placed in a separate species, it is not known if the pygmy chimp is just a scaled down version of the common chimp, a phyletic dwarf, or if it is distinct enough to be placed in a separate genus-as blood group serology studies suggest. [27]
If relationship questions like these cannot be solved regarding primates which are living and where many complete skeletons are available for study, we have every right to challenge the claim that Australopithecus ramidus is the 'missing link' proving human evolution, all based on 17 fossil fragments.
Anthropologist Adrienne Zihlman (University of California, Santa Cruz) has long emphasized the remarkable similarity between the skeleton of the pygmy chimp and that of our alleged early ancestors.
“Except for the pelvis the P. paniscus skeleton shows a striking resemblance to fossils of the earliest hominid Australopithecus.” [28]
Her illustration showing the left half of the skeleton of a pygmy chimp joined to the right half of the skeleton of the fossil Lucy (Australopithecus afarensis), revealing the close match, has appeared in a number of science periodicals. [29], [30], 31 Zihlman, of course, does not believe that Lucy was a pygmy chimp. She feels that the pygmy chimp is the 'living link' to man's earliest ancestor—that the common ancestor must have looked something like a pygmy chimp. Other anthropologists such as Vincent Sarich (University of California, Berkeley) and Henry McHenry (University of California, Davis) tend to share her view.
Zihlman's view implies that in the course of the pygmy chimp's evolutionary history it has remained quite similar to the 'ape-human' ancestor, while the common chimpanzee and the hominids have undergone considerable morphological change.
What is the reason given for this remarkable disparity? It is that the pygmy chimp represents the more ‘primitive’ condition, having gone through fewer specialized adaptations. At one time, it is suggested, pygmy chimps were far more widespread in Africa. Some of the roving chimp bands found themselves in totally new environments and eventually turned into new species. But stragglers who remained in the central African rain-forest were perfectly suited to their habitat, and so the pygmy chimps remain today much as they have been for four million years. With this type of speculation, it is easy to understand why evolution has been termed “a fairy-tale for adults.”
The Bipedalism Problem
In spite of the universally assumed bipedality of the early hominids, the bipedality of those creatures is far from certain. Carol V. Ward (University of Missouri, Columbia) writes:
The origin of hominid bipedality is one of the most controversial issues paleoanthropology. [32]
Bernard Wood agrees:
Bipedalism is a fundamental human characteristic yet virtually nothing is known about its origins. [33]
Citing the research of K. D. Hunt (University of Michigan), Wood makes what seems to be a valid distinction between bipedal posture and bipedal locomotion. Observations of chimpanzees in their natural habitat reveal that 80 percent of their bipedalism was postural and related to feeding. Only four per cent of their bipedalism was related to locomotion, and that was a rather ungainly shuffling between feeding sites. Hunt argues that early hominids, such as Lucy, were postural bipeds but not locomotional bipeds, and that locomotional bipedality came only with the appearance of Homo. [34] Evolutionists have not taken this distinction into consideration when arguing for the bipedality of early hominids. It suggests that 'early hominid' bipedality has no relationship to human bipedality.
Pygmy chimps, like common chimps, are basically quadrupedal knuckle-walkers when on the ground. However, observations of them in the wild and in captivity reveal that they walk bipedally about 10 per cent of the time both on the ground and in the trees, especially when they are carrying objects or engaged in behavioral displays. The ability of primates like chimpanzees to be bipedal under certain situations may be what evolutionists are mistaking for an alleged evolutionary trend from quadrupedal to bipedal locomotion.
Evolutionists claim that no fossils of chimpanzees have ever been found. The evidence suggests that fossils of chimpanzees have been found, but the blinding power of a naturalistic evolutionary philosophy, and the determination of evolutionists to find evidence for it has not allowed these fossil chimpanzees to be recognized for what they are.
Recent Trends Regarding Homo Habilis
The demise of Homo habilis has been amazingly swift. Although controversy has always surrounded the taxon, it has nevertheless been almost universally regarded as the transition between the australopithecines (which everyone acknowledges were non-human) and Homo erectus (which virtually everyone acknowledges was fully human). Over 100 numerically designated fossils or fossil assemblages have been formally or informally allocated to Homo habilis or have been declared to have affinities with that hypodigm., (a term used to refer to all of the fossils allotted to the species under consideration). Homo habilis thus represents an extensive fossil collection.
However, the discovery of Olduvai Hominid 62 in Tanzania in 1986 brought out in bold relief some of the nagging questions that had plagued Homo habilis. Those questions were:
The problem of the large range of cranial sizes in the taxon. Whereas some workers explained this in terms of sexual dimorphism, others felt that the size range exceeded that which could reasonably be explained by calling the larger ones males and the smaller ones females.
The problem of the large range of morphological variation in the post-cranial material, some of it being juvenile material which is difficult to diagnose. Louis Leakey had explained some of this range of morphology as evolutionary change over time. Others felt that this morphological variation exceeded that which would be expected within a single species.
The problem of reversals. Most workers who accepted Homo habilis as a legitimate taxon also accepted the Homo erectus fossils as human ancestors. Thus, to go from the thin-walled and high-domed cranium of Homo habilis to the thick-walled and low-domed cranium of Homo erectus and then back to the thin-walled and high-domed cranium of modern humans represented reversals in both cranium thickness and cranium morphology. Reversals are not supposed to happen in an evolutionary sequence.
These problems, and others, have caused a major shift in attitudes toward Homo habilis. It is safe to say that a large majority of palaeoanthropologists now feel that the Homo habilis hypodigm represents at least two, if not three, different species that have mistakenly been lumped together Confusion is now the rule. Richard Leakey states that of all the fossils considered to belong to the Homo habilis hypodigm,
…at least half probably don't. But there is no consensus as to which fifty percent should be excluded. [35]
There are three major, but tentative, scenarios involving Homo habilis. The first one is suggested by Donald Johanson and Tim White. It is the least likely one, but it has the advantage of salvaging their 1979 arrangement of the hominid family tree in which Homo habilis was the transition between the australopithecines and Homo erectus. They propose that the entire Homo habilis hypodigm represents just one species. The differences in morphology and size among the fossils are the result of evolution over time (early habilis versus late habilis) plus sexual dimorphism and geographic factors.
A second interpretation is proposed by Richard Leakey and Alan Walker (Johns Hopkins University). They suggest that the Homo habilis hypodigm consists of two species—one Homo and the other australopithecine. The true Homo habilis would consist of the larger-toothed, bigger-brained, and presumably bigger-bodied component of the hypodigm. This species would include fossils such as KNM-ER 1470, 1481, 1390, and Olduvai Hominids 7 and 16. They consider the smaller fossils of the hypodigm to be just varieties of Australopithecus africanus—late survivors that were contemporary first with Homo habilis and later with Homo erectus. The fossils in this second category would include KNM-ER 1803, 1813, and Olduvai Hominids 13 and 24. [36]
A third possibility is suggested by Bernard Wood. He also believes that there are two species involved in the Homo habilis hypodigm—in this case, two Homo species. Whereas Leakey and Walker believe that the larger fossils represent the true Homo habilis, Wood believes that the smaller fossils In the hypodigm represent the true Homo habilis. He proposes a new Homo species for the larger fossils. [37] There are other workers who speculate that as many as three different species make up the Homo habilis hypodigm. It seems clear that Homo habilis is a phantom species, and that the fossils labeled habilis belong to other categories.
In spite of the crisis, it is obvious that evolutionists need a transition between the australopithecines and Homo erectus. Thus, there is much effort to try to salvage at least a part of the Homo habilis hypodigm. The seriousness of the situation is seen in this confession by Milford Wolpoff (University of Michigan):
…the phylogenetic outlook suggests that if there weren't a Homo habilis we would have to invent one… [38]
The history of Homo habilis suggests that this is indeed what has happened.
Recent work on the semi-circular canals (bony structures which in life housed the organs of balance) in a range of fossil skulls is pertinent. Not only did all australopithecine specimens tested show that these creatures did not walk habitually upright (whereas all the erectus specimens did); when a so-called Homo habilis specimen was tested, it was shown to be even less suited to upright walking than both australopithecines and modern-day great apes. [39], [40], [41]
RECENT TRENDS REGARDING HOMO ERECTUS
With Homo habilis in disarray, the taxon Homo erectus takes on greater importance in the crucial transition from the australopithecines to the archaic Homo sapiens and Neanderthal fossils. Yet, the more than 222 Homo erectus fossil individuals which comprise the taxon have stubbornly resisted efforts to place the taxon into a neat evolutionary continuum. Evolutionists face a number of problems regarding Homo erectus) most of which are unknown except to researchers in this field. These problems include:
the origin of Homo erectus,
its fate, and
the lack of evolutionary change within the taxon over more than one million years of alleged evolutionary-history.
Obviously, the turmoil regarding Homo habilis has profound effects on the problem of the origin of Homo erectus, and the three positions on the matter are related to the three attitudes regarding Homo habilis. The first position on the origin of Homo erectus is that of Donald Johanson and Tim White. Based upon their evaluation of Olduvai Hominid 62 and the rest of the Homo habilis hypodigm, they feel that the entire hypodigm constitutes a single species which is ancestral to Homo erectus. [42] Utilizing the punctuated equilibria model of evolution, they propose that the three foot tall Homo habilis species at 1.8 million years ago evolved into Homo erectus by about 1.6 million years ago.
Johanson and White's explanation for the origin of Homo erectus fails for at least three reasons. First, the relatively complete Homo erectus skeletons KNM WT 13000 and KNM-ER 1808 both come from just that 1.6 million years ago time-frame. They reveal that at that time on the evolutionist time-scale, Homo erectus was up to six feet tall. For a species to double in size in such a short time is asking too much even of the punctuated equilibria model.
Second, Homo erectus couldn't have evolved from Homo habilis in the 1.8 to 1.6 million years ago time-frame because Homo erectus was already on the scene, possibly as early as 2.0 million years ago. G. Philip Rightmire (State University of New York, Binghamton) states that skulls and postcranial fossils of Homo erectus have been found in several areas near Koobi Fora, Lake Turkana, Kenya, and that: 'The earliest of these fossils must be older than 1.6 million and perhaps as old as 2.0 million years…' [43]
Third, as was mentioned earlier, to go from Homo habilis to Homo erectus to Homo sapiens represents reversals in both cranial wall thickness and skull morphology. This would imply that if Homo erectus is in the mainstream of human evolution, Homo habilis cannot be; whereas if Homo habilis is in the mainstream, Homo erectus is excluded. In either case, it compromises the idea that Homo erectus evolved from Homo habilis.
Richard Leakey and Alan Walker have another proposal for the origin of Homo erectus. Believing that the Homo habilis hypodigm involves two species, they feel that the larger fossils of that hypodigm are the true Homo habilis and the true ancestors of Homo erectus. However, this scheme also fails because of the reversal problem. Leakey himself claims that there is a built-in mechanism in the evolutionary process that eliminates reversals. [44]
Bernard Wood has still another solution for the origin of Homo erectus. He believes that the Homo habilis hypodigm involves two Homo species, with the smaller fossils of that hypodigm representing the true Homo Habilis. This would include all of the Olduvai Gorge material as well as the smaller fossils from Koobi Fora, Lake Turkana, Kenya.
He proposes a new taxon for the larger fossils at Koobi Fora, Homo rudolfensis (after Lake Rudolf, the old name for Lake Turkana). The fossils in this new taxon include KNM-ER 1470, 1472, 1481, 1590, 1802, and 3732.
Wood complicates matters further by proposing another new taxon, Homo ergaster made up of fossils KNM-ER 3733, 3883, and KNM-WT 15000, which virtually every other investigator classifies as full-fledged Homo erectus. Wood calls these three fossils African precursors of Homo erectus. [45] Wood's interpretation thus places three Homo species in the 1.5 to 2.0 million years ago time-frame, with Homo erectus evolving from Homo ergaster. But in his phylogenetic chart, Wood has Homo ergaster, Homo habilis, and Homo rudolfensis arranged as sister species with a question mark for the origin of all three of them. Thus, in Wood's view, Homo erectus comes from Homo ergaster, with the origin of Homo ergaster unknown.
In contrast to popular belief, the origin of the fully human taxon called Homo erectus is not known. Among evolutionists, anything is possible. Homo erectus might come from the entire Homo habilis hypodigm (Johanson and White), from the larger fossils of that hypodigm (Leakey and Walker), or from unknown fossils which may or may not be of that hypodigm (Wood and others). Richard Leakey states that the human evolution story at 2.0 million years ago is unclear and he hopes that new fossils will cast light on the subject. [46]
G. Philip Rightmire reveals the naked truth
Just how Homo erectus first evolved is one of the major issues in Paleoanthropology. Here the fossils and the stratigraphic record are limited and many details may never be resolved. [47]
Homo erectus is in fact a false category. Evolutionists have taken true human ancestors and tried to make them evolutionary ancestors. The somewhat different skull morphology of Homo erectus may be the result of the environment of the post-Flood Ice Age. [48] Since Homo erectus did not evolve but was a true member of the human family created by God in His image, it is not surprising that evolutionists have great difficulty in finding an evolutionary ancestor for him.
Lack of Evolution Within Homo Erectus
It is common for evolutionists to claim documentation for evolutionary change within the Homo erectus taxon based primarily on the fossils from Java. Many natural history and anthropology museums have such displays, including the San Diego Museum of Man. In fact, if evolution were true, change within this taxon is exactly what we would expect, and it is what evolutionists say should happen. However, that claim involving the Javanese fossils is invalid for two reasons.
First, the stratigraphy of the Javanese fossil beds was still unclear long after those fossils were discovered. Second, very few of the fossils from Java were seen in situ by the investigators who 'discovered' them, such as Dubois and von Koenigswald. The bulk of the Javanese fossils were uncovered by hired nationals who knew nothing about geology or stratigraphy, with Dubois or von Koenigswald arriving on the scene well after the fact. All palaeoanthropologists know that the Javanese fossil dates are uncertain. Those displays showing evolution within the Javanese Homo erectus fossils are raw propaganda designed to influence the unknowing public toward evolution.
There is no significant change throughout the history of Homo erectus that would in any wise document evolution. That is true not only of the Javanese fossils but of the entire taxon. The leading investigator in this area, G. Philip Rightmire, has evaluated specific characteristics of Homo erectus over its alleged million year history, comparing early erectus with late erectus. He concludes that there is no characteristic that changes in a significant evolutionary way. 'Following the emergence of Homo erectus, systematic change is not easily documented.' [49]
Commenting on the many regions of the world inhabited by Homo erectus, he adds:
Populations inhabiting these far flung regions of the Old World are anatomically similar, and the morphology of the species seems to have changed little over more than a million years. [50]
The Fate of Homo Erectus
In the past ten years, there has been a shift in emphasis in the study of human evolution—from the origin of all humans to the origin of modern humans. Central to the question of the origin of modern humans is the fate of Homo erectus. The two views that address this issue are known as:
the “Out of Africa,” “Noah’s Ark,” “African Eve,” or “Mitochondrial Eve” theory, and
the Regional Continuity Model.
One of the prime movers of the 'African Eve' model is Christopher Stringer (British Museum-Natural History). The leading advocate of the Regional Continuity Model is Milford Wolpoff.
It is unfortunate that some creationists have seized on the 'African Eve' concept as if it were pointing inevitably to the Eve of the Bible. While it is true that biblical considerations mean that the finding of one mitochondrial 'signature' in all females would be a consistent expectation, there is no necessary relationship, between the two. Even Richard Leakey is concerned that this alleged 'mother of us all' was dubbed Eve, because the theory does not deal with a lone mother, but one woman in a population of thousands. It is just that her mitochondrial DNA is the only one that has been passed down via the female line.
The 'African Eve' theory holds that modern humans evolved only in sub-Saharan Africa, possibly from some kind of African Homo erectus stock, about 200,000 years ago. About 100,000 years ago a portion of that population migrated out of Africa into the Near East and then into both Europe and Asia. Whatever primitive peoples were inhabiting those areas, specifically Homo erectus, were exterminated by the invading forces without any genetic mixing. Thus, all modern populations are genetically related to one woman who was in this migrating African group of modern humans.
Kenneth Kennedy (Cornell University) points out that the 'African Eve' model is very 'politically correct'. [51] In a world where males are accused of domination, it is comforting to know that a woman is the hero of the plot. Without her, we would not exist. With feelings of exploitation being held by non-Western peoples, a victorious non-Western minority overcoming the world is also welcome news. Further, a sapiens species out of Africa emphasizes the unity and brotherhood of all modern peoples in spite of external differences. Those holding to punctuated equilibria also favor the 'Out of Africa' model as a textbook illustration of abrupt evolutionary change which, among other things, eliminates the Neanderthals from the modern human lineage. The 'African Eve' model even has a hint of catastrophism in it, which is coming into favor in some evolutionist circles.
The “African Eve” model became immensely popular because biochemists claimed strong scientific, namely molecular, evidence for it. However, the model has recently fallen on hard times. The alleged scientific evidence for it was based upon an improper interpretation of results from a complex computer program. One of the initial investigators, Mark Stoneking (now at Pennsylvania State University), admits that the results indicating an African origin are flawed. [52]
The Asian fossil record clearly falsifies the 'Out of Africa' model. At least 67 fossil individuals having a Homo erectus morphology are dated more recently than 100,000 years ago. (All of this uses evolutionary dating for the sake of the argument of course.) Thus, Homo erectus was not exterminated by more modern invaders, but persisted until very recent times. Evolutionists have concealed this fact by calling these fossil individuals Homo sapiens, based only on the late dates of the fossils, disregarding their morphological similarity to Homo erectus. 1 have detailed these matters elsewhere. [53]
The other major model dealing with the fate of Homo erectus and the origin of modern humans is the Regional Continuity Model. This model assumes the presence of Homo erectus populations in Europe, eastern Asia, and Africa. It then holds that modern Europeans evolved from the Homo erectus population In Europe, modern Africans evolved from the Homo erectus population in Africa, and modern Chinese evolved from the Homo erectus population in eastern Asia.
However, there is a problem. A great unsolved mystery is that all humans are of one species—Homo sapiens. Our world-wide interfertility is unique in the biological world. The 'Out of Africa' model sought to solve the problem by having all modern humans descend from one small African population. However, if modern populations evolved independently in different parts of the world, especially from a different species, Homo erectus, how does the Regional Continuity Model explain this world-wide interfertility? It does so by postulating, without any evidence, a large exchange of genes throughout the world and throughout all of human evolutionary history. A world-wide flow of genes to maintain the solidarity of the human species is absolutely essential to the model. Milford Wolpoff states:
…this happened to some extent because all these populations were interconnected by a flow of genes. People were coming and going, exchanging wives and so on. We think all humanity was interconnected this way. [54]
This Regional Continuity Model, unfortunately, seems to be based upon genetic fantasy. Geneticist Shaham Rouhani (University College, London) observes:
Even under ecologically identical conditions which rarely exist in nature, geographically isolated populations will diverge from each other and eventually become reproductively isolated… It seems to me that the multiregional model of modern human origins is therefore theoretically implausible. [55]
Doubts are also expressed by geneticist Luigi Luca Cavalli Sforza (Stanford University):
Very large populations have a genetic inertia… It would take a very long time for mutations to move through such a population. I don't see how the multiregional model could work [56]
The Regional Continuity Model does not find adequate support in the fossil record either. Most researchers feel that there are no unequivocally recognized specimens of Homo erectus in Europe. [57], [58] Thus, as far as we know, there was no population of Homo erectus for modern Europeans to evolve from. Asia presents problems also. Rightmire comments:
…there is not much basis for postulating direct' evolutionary continuity from Homo erectus to Homo sapiens in the Far East, either in the north or in Australasia. [59]
Only Africa is left. Regarding Africa, W. W. Howells (Harvard University), referring to the work of Rightmire makes a most amazing statement:
The persisting features are such that, in Africa, a transition from Homo erectus to Homo sapiens should be accepted, taking place in a region not determined as yet. [60] (emphasis added)
Is that fact, or is that faith?
Regarding the fate of Homo erectus and the origin of modern humans, both the 'Out of Africa' model and the Regional Continuity Model are contradicted by the human fossil record. There is simply no viable scientific explanation for the fate of Homo erectus or the origin of modern humans in an evolutionary model.
When humans reject the authority of God's Word and the historicity of the Genesis account of human creation, it should not surprise us that more than 100 years of secular research into human origins has yielded no reliable answers.
Author: Dr. Marvin L. Lubenow, “Paleoanthropology in Review,” CEN Technical Journal, Vol. 10, No. 1 (1996), pp.10-17. Supplied by Creation Ministries International.
ABOUT THE AUTHOR: Dr. Marvin L Lubenow received his doctorate from Christian Heritage College, El Cajon, California, where he is a Professor Emeritus. He received his M.Th. from Dallas Theological Seminary and a M.S. in anthropology and paleontology from Eastern Michigan University. He is the author of Bones of Contention, the product of more than 25 years of research in paleoanthropology.
Copyright © 1996, 1997, 2000, Creation Ministries International, All Rights Reserved—except as noted on attached “Usage and Copyright” page that grants ChristianAnswers.Net users generous rights for putting this page to work in their homes, personal witnessing, churches and schools. / HTML coding: Films for Christ and David M. Harris (volunteer)
References
- Gibbons, A, 1992. Sorting the hominoid bone pile. Science, 156:176. Dr Meave Leakey, Richard Leakey's wife, recently announced that an ape-like fossil maxilla with four teeth, discovered in Kenya some time ago, had recently been related to between 24 and 27 million years ago. This redating would place this fossil in the hominoid gap. Return to text
- White, T.D., Suwa, G. and Asfaw, B., 1994. Australopithecus ramidus a new species of early hominid from Aramis, Ethiopia: Nature, 371:306-312. Return to text
- White, T. D., Suwa, G. and Asfaw, B., 1995. Corrigendum: Australopithecus ramidus new species of early hominid from Aramis, Ethiopia. Nature, 375:88. Return to text
- WoldeGabriel, G., White, T. D., Suwa, G., Renne, P., de Heinzelin, J., Hart, W. K. and Helken, G., 1994. Ecological and temporal placement of early Pliocene hominids at Aramis, Ethiopia. Nature, 371:330-333. Return to text
- White et al., Ref. 3, p. 88. Return to text
- White et al., Ref. 2, p. 306. Return to text
- Groves, C., 1995. Gift from Aramis 'the missing link'. The Canberra Times, April 11,1995, p. 7. Return to text
- Time October 3, 1994, p. 68. Return to text
- Newsweek October 3, 1994, p. 57. Return to text
- Groves, Ref. 7. Return to text
- Ref. 8, p. 69. Return to text
- Ref. 9, p. 56. Return to text
- White et al., Ref. 2, p. 306. Return to text
- Wolde Gabriel et al., Ref. 4. Return to text
- Kappelman, 3. and Beagle, 3., 1995. Age of early hominids. Nature, 376:558-559 Return to text
- Kappelman and Fleagle, Ref. 15, p. 559. Return to text
- Lubenow, M.L., 1991 Bones of Contention, Baker Book House, Grand Rapids, Michigan, pp. 247-266. Return to text
- Wood, B., 1994. The oldest hominid yet. Nature, 371:280. Return to text
- White et al., Ref. 2, p.310. Return to text
- Fischman, 3., 1994. Putting our oldest ancestors in their proper place. Science, 265:2011. Return to text
- Lawrence, M., 1985. Significance of enamel-thickness in hominoid evolution, Nature, 314:260-263. Return to text
- Grine, F., 1991 Variability in enamel thickness and structure among molar tooth classes in Homo sapiens. American journal of Physical Anthropology, Supplement 14, p. 85. Return to text
- White et al., Ref. 2, p. 306. Return to text
- White et al., Ref. 2, p. 310. Return to text
- White et al., Ref. 2, p. 312. Return to text
- Gee, H., 1995. Uprooting the human family tree. Nature, 373:15. Return to text
- Leutenegger W., 1985. Pan paniscus. Science; 228:982. Return to text
- Zihlman, A., 1992. The promiscuous primate. Nature, 359:786. Return to text
- New Scientist, 15 November, 1984, p. 45. Return to text
- Science 83, June 1983, p. 45. Return to text
- Science News, 123 (March 19, 1983):92. Return to text
- Ward, C., 1992. Book review. American Journal of Physical Anthropology, 90(3): 387. Return to text
- Wood, 13., 198?. Four legs good, two legs better, Nature, 363:587. Return to text
- Ward, Ref 32. p 388 Return to text
- Leakey, R. and Lewin R., 1992. Origins Reconsidered, Doubleday, New York, p. 132. Return to text
- Leakey and Lewin, Ref. 35, pp. 97-120. Return to text
- Wood, 13., 1991 Origin and evolution of the genus Homo. Nature, 355:783-790 Return to text
- Wolpoff, M., 1992. Book Review. American Journal of Physical Anthropology, 89(3): 401 Return to text
- Wieland, 0,1994. New evidence: only people ever walked really upright. CEN Technical Journal, 8(2): 127-128. Return to text
- Shipman, P., 1994. Those ears were made for walking. New Scientist, 143(1936):26-29. Return to text
- Spoor, F., Wood B. and Zonneveld, F., 1994. Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature, 369:645-648 Return to text
- Johanson, D. Masao, K, Eck, G., White T., Walter, R., Kimbel, W., Asfaw, 13., Manega, P., Ndessokia, P. and Suwa, G., 1987. New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature, 327:205-209. Return to text
- Rightmire, G. P., 1990. The Evolution of Homo erectus, Cambridge University Press, Cambridge, p. 86. Return to text
- Leakey and Lewin, Ref. 35, p. 297. Return to text
- Wood, Ref. 37. Return to text
- Leakey and Lewin, Ref. 35, p. 33. Return to text
- Rightmire, Ref. 43, p. 9. Return to text
- Lubenow, Ref. 17, pp. 141-156. Return to text
- Rightmire, Ref. 43, p. 202. Return to text
- Rightmire, Ref.: 43, p. 204. Return to text
- Kennedy, K., 1992. Book Review. American Journal of Physical Anthropology, 89(2):272. Return to text
- Hedges, S. B., Kumar S., Tamura, and Stoneking, M., 1992. Human origins and analysis of mitochondrial DNA sequences. Science, 255:737-739. Return to text
- Lubenow, Ref. 17, pp. 120-133 and 144-156. Return to text
- Putman, J. J., 1988. The search for modern humans. National Geographic, 174(4):463. Return to text
- Leakey and Lewin, Ref. 35, pp. 213-214. Return to text
- Leakey and Lewin, Ref. 35, p. 214. Return to text
- Howells, W. W., 1980. Homo erectus—who, when and where: a survey. Yearbook of Physical Anthropology, 23, Alan R. Liss, Inc., New York pp. 12, 13 and 15. Return to text
- Leakey and Lewin, Ref. 35, p. 209. Return to text
- Rightmire, Ref. 43, p.233. Return to text
- Howells, Ref.: 57, pp. 1617 Return to text