What are the Genesis “kinds”?
Baraminology—classification of created organisms

For decades creationists have been using the word "kind," “type,” or “group” for their envisioned categories of genetically unrelated organisms, including all those formed by the Creator during the Creation Week. Within each of these categories the various species, subspecies, and varieties were conceived to have diversified from common ancestral stock. However, until recent years there has not been a serious comprehensive methodology of classification focusing on characterizing each original category, which is separated by genetic gaps from all other categories. Now baraminology (with discontinuity systematics) has developed into a fruitful approach to classification within the creation model. Terminology and methodology have been developed, and the first scientific baraminology conference was held in the summer of 1999.
[Editor's note: The term baraminology means study of created kinds. It was coined from the Hebrew words "bara" (created) and "min" (kind).]
Introduction
Basic human attributes include classification and identification. People do these so constantly that the practices are essential to our way of living. Individuals have learned the classification of thoughts and things, and as they interact with their environments they persistently identify cognitions as smells, sights (sky, food, animal, plant, etc.), sounds (honk of horn, bark of dog, music, etc.), savors (sweet, sour, etc.), and other sensations such as rough and smooth. The complexity of all this is astounding.
Interestingly, the Bible includes an account of God’s bringing to Adam all the terrestrial animals and birds (Genesis 2:19) so that Adam could observe and name them. This story may have a deeper significance than a cursory reading would reveal. In the process of naming the creatures Adam learned something about himself. He realized his capacity to perceive and to discover patterns. Some critters wore scales and others hair or feathers.
In addition, Adam would learn something about God--the Designer. There in the creation was an expression of the Lord’s handiwork. After Adam discovered nature he never would be the same again. The stars, the animals, the plants--all were part of God’s creation. The stage now was set, and subsequently billions of science students would play their part upon the stage of life and during that time sense the excitement of discovering God’s message in nature (Psalm 19:1; Romans 1:19,20; and see ReMine, 1993).
Taxonomy and Systematics
Pondering these matters makes it easier for us to realize the basic importance of classification in biological studies of extinct and extant forms of life. Taxonomy is the term used for the science of classifying living things according to their natural groupings. Essentially, scientists of today utilize a taxonomic system introduced by the Swedish botanist, Linnaeus, about 250 years ago.
However, there are different approaches used by scientists for studying patterns of life and for classification. These are called systems of taxonomy and they lie under the general heading of systematics. For studies of nature’s patterns ReMine (1993, p. 444) has compared the diverse procedures to the functions of different types of film (infrared, ultraviolet, and x-ray film). For example there is phyletic or evolutionary systematics (or Darwinian systematics) in which macroevolution (involving large changes) is assumed. See Figure 1.
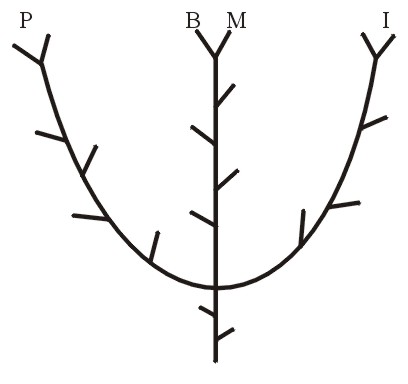
Figure 1.
Systematic schemes which place less stress upon evolution may be termed phenetic because they are based upon appearances of features of the organisms and not necessarily their possible large scale evolutionary relationships. Also there is the cladistic approach which focuses on common or so-called "shared/derived" characters. Most importantly, popular methodology in phyletic, phenetic and cladistic procedures all have been adapted to macroevolutionary theorizing.
The earliest methods were phenetic which conferred the distinct advantage of being objectively observable and most verifiable. Critics of basic phenetics feel that its simplicity does not encourage adequate consideration of historical causation. Nevertheless, those who employ baraminology (with discontinuity systematics) as presented in this paper, can be agnostic regarding popular phenetic and cladistic methodology, but at least they do employ some phenetic methodology as one of their tools.
Discontinuity Systematics and Baraminology
Scientists who have preferred something other than a macroevolutionary framework recently have developed what they believe to be a more realistic systematics based upon the discontinuities or typology found in nature. This methodology appropriately has been termed discontinuity systematics (formally presented by Walter J. ReMine, 1990), or when combined with Biblical revelation, baraminology (a term introduced by Kurt P. Wise, 1990). Baraminology may be defined as a taxonomy based upon the created kinds (see Bartz, 1991; Frair, 1991; 1999; and Figure 2).


Figure 2.
The word “baramin” was conceived by Frank L. Marsh and first published in 1941; it is derived from the Hebrew verb bara, create and min, kind (also see Marsh, 1969; Williams, 1997).
Since classification underlies all biological investigations, it is quite significant that creationists now have an active focus on this topic. Substantial progress has been made since 1990, and baraminologists have developed their own terminology which at this time appears to be quite practical for those doing systematic research. The major purpose of baraminology is to determine which organisms share common ancestry.
Marsh employed the term baramin in an inclusive way for an entire group of known, unknown, and possibly inferred organisms sharing genetic relationship. But now the focus is more specific, and only those specimens which can be studied as living or extinct (including fossil) specimens may be included in the current four main baraminic groups. The terms employed as the four primary baraminic categories are holobaramin, monobaramin, apobaramin, and polybaramin.
Holobaramins
In baraminology the primary term is holobaramin from the Greek holos for whole. The holobaramin is all and only those known living and/or extinct forms of life understood to share genetic relationship. It is an entire group believed to be related by common ancestry.
So now each natural group of related plants or of related animals constitutes a holobaramin; or in more specific creationist terminology the holobaramin consists of all known organisms in a group beginning after God created the original organisms (see Wise, 1992). The holobaramin may be represented as a branching tree, the nodes and tips of the branches representing all the known members (subspecies, species, etc.) of the “kind” (“group”, or “type”). See Figure 2. When individuals or groups of apparently related specimens are being compared they may be designated as holobaraminic if they constitute parts of one holobaramin.
During recent past decades the creationist researchers have employed the terms “kind”, “group” and “type” generally interchangeably; or as individuals the researchers have preferred one or another particular general name for Marsh’s “baramin,” and also for what more specifically and currently may be designated as holobaramins. Now systematists of particular taxons of plants or animals may discard the older terminology and construct their trees showing holobaraminic affinities, and thus the boundaries of common descent.
An important example of a holobaramin would be humans, Homo sapiens. At the tips of the holobaraminic branches are the various races (Caucasians, Ethiopians, Mongolians, Amerindians [Amerinds or Native Americans], etc.). See Figure 3. A member of any of these races potentially would be inter-fertile with a spouse of the opposite sex from any other race.
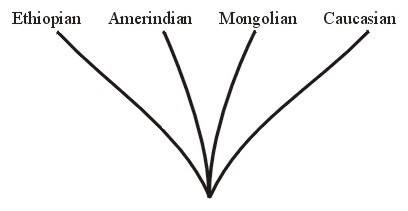
Figure 3.
It is not uncommon to find in the anthropological literature reference to upward of eight human geographical races with even additional intermediate populations. However, it is not my intention in this article to enter into discussions of the different options for what is expressed here in the text or in any of the figures, but merely to illustrate the taxonomic principles involved.
Another holobaramin could consist of the sea turtles (see Wise, 1992; Robinson, 1997). A diagram showing general forms of living and fossil sea turtles may be found in Lutz and Musick, 1997, p. 8. This diagram is called a “cladogram” and is based upon studies by specialists Gaffney and Meylan, but not all authorities agree with their assessment of available data. See also Hirayama, 1998. Figure 4 is a very generalized representation for all living and extinct marine turtles.
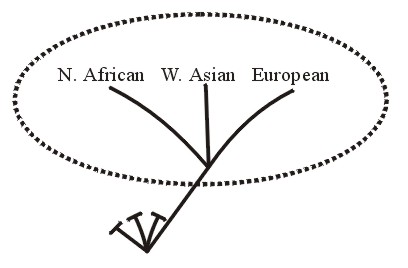
Figure 4.
In all of these types of studies the actual goal of discontinuity systematics is by means of empirical evidence to determine the boundaries of common descent and thus to converge on the holobaramins.
The different members of a holobaramin could have resulted from a sorting out to the offspring of different genes (DNA) from parental organisms. This is a common occurrence today. Or, since the time of creation there could have been some hereditary modifications of the DNA (mutations), and these were passed on to the diverging offspring. Selection in nature could have influenced the potential for survival of the diverse siblings.
Monobaramins
The second term used in baraminology is monobaramin (mono, from the Greek for single or one). The term monobaramin is defined by ReMine (1993, p. 444) as:
a group containing only organisms related by common descent, but not necessarily all of them. (A group comprising one entire holobaramin or a portion thereof).
When a holobaramin is represented by a tree, one or more branches of that tree would be a monobaramin. For example, among humans, the caucasians would be a monobaramin (Figure 5).
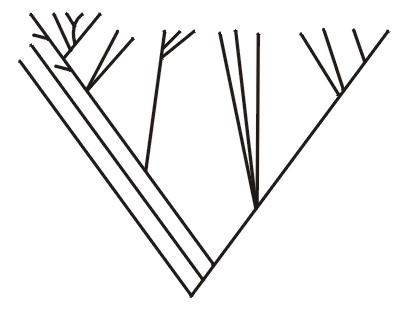
Figure 5.
Or for the sea turtles, the five current types living in oceans around the world constitute a monobaramin (Figure 6A from Frair, 1982; and see Iverson, 1992, p. 80). Also, the group of green turtles, Chelonia, or the branch containing the ridley turtle, Lepidochelys, each would be a monobaramin (Figure 6B). Individuals or groups may be referred to as monobaraminic if they represent parts of a holobaramin (Figures 5, 6A, 6B, and 7B).
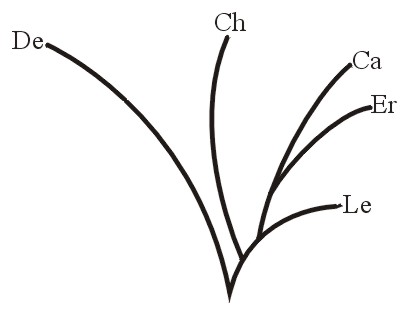
Figure 6a.
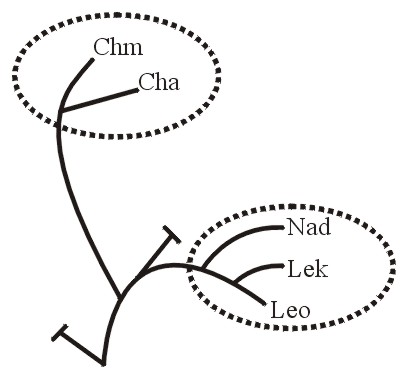
Figure 6b.
In addition, systematic studies on particular monobaramins where there has been diversification (as the appearance of more recognizable species, subspecies, varieties, etc.) the research on a monobaramin would not differ essentially depending on the systematic philosophy of the investigators. The difference between a (1) phyletic, in the sense of a Darwinian macroevolutionary perspective, and a (2) baraminic (creationist, “limited change”, or microevolutionary) discontinuity systematics viewpoint mainly would be that the former involves the use of empirical data for extrapolating to some perceived earlier ancestors. But the baraminologist maintains that thinking about phylogeny should not extend beyond convincing evidence, and that scientists should be relieved of their sense of obligation philosophically to construct extensive phylogenies (evolutionary trees) in the absence of compelling facts.
Apobaramins
A third baraminic term is apobaramin (Greek apo, away from), which "is a group consisting of the entirety of at least one holobaramin" (Wise, 1999-2000). It may contain a single holobaramin or more than one holobaramins. "But it must contain the entirety of each of the one or more holobaramins within it". No member organism of a holobaramin within an apobaramin shares ancestry with any organism outside of its own holobaramin (this being based upon the definition of holobaramin). See Figure 7A.
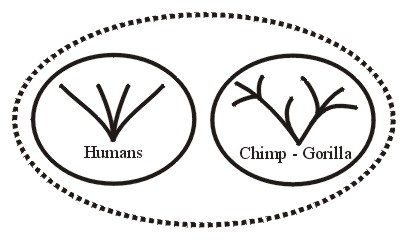
Figure 7a.
The adjective apobaraminic refers to the association between or among distinctly unrelated groups (holobaramins). For example all humans as a group would be apobaraminic because none of its members shares ancestry with any other organisms. The group of all humans and all turtles also would be apobaraminic because no human or turtle shares ancestry with any non-human or non-turtle organisms.

It is believed that the horses (horses, donkeys, and zebras) all are related because they can hybridize, and therefore they belong to a holobaramin. Additionally there is a “dog” holobaramin with monobaraminic branches for the wolves, another for the hyenas, another for the coyotes, for jackals, and more for the hundreds of pet-dog breeds. “Cats” constitute another holobaramin with monobaraminic branches for the lion and the tiger, for the pumas, another for the lynx, domestic cats, etc. (see O’Brien, 1997). A group of all the horses (equids), all the dogs (canids), and all the cats (felids) would be apobaraminic because no horse or dog or cat shares a genetic relationship with any organism which is not a horse, a dog, or a cat.

The turtle apobaramin may consist of one, two, three or four holobaramins (see Wise, 1992). In this present article I am considering the sea turtles to constitute a holobaramin; therefore a group containing the sea turtle holobaramin, all equids, all canids, and all felids would be apobaraminic because none of the members of any of these four holobaramins shares genetic relationship with any specimens outside their respective holobaramins.
The term apobaramin is a term useful especially during evaluations of two types of organisms (pairwise comparisons). Utilizing pairwise comparisons is the most common taxonomic procedure.
For example, the current Order Primates includes apes, humans, lemurs, monkeys and tarsiers. All races of humans belong to one holobaramin; whereas chimpanzees (chimps) along with gorillas are members of another holobaramin.

So a group containing the human holobaramin and the chimp-gorilla holobaramin would be apobaraminic (see Figure 7A). Further, a collection of the human holobaramin with any or all the other primate holobaramins would be apobaraminic. No member of any of these holobaramins would share any ancestry with a member of any of the other holobaramins within or even outside this apobaramin. See Robinson and Cavanaugh, 1998a for a baraminic study of Primates.
Conclusions regarding a holobaraminic chimp-gorilla relationship (Figures 7A and 7B) are based upon Hartwig-Scherer, 1998.
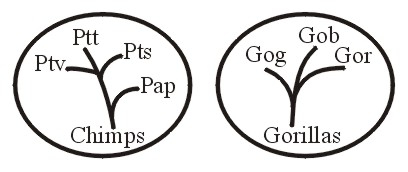
Figure 7b.
Also Cavanaugh (1999-2000) has informed me that a restudy of data from Robinson and Cavanaugh (1998a) supports the holobaraminic status of chimps and gorillas. However, as explained by Klein (1999, pp. 135-136) the fossil record provides very little that is of any use in understanding the history of chimps and gorillas. When fossils convincingly have been determined to be related to chimps and gorillas they should be added to their holobaramin. Even possibly the chimp-gorilla group should be divided into two separate holobaramins. Figure 7A illustrates how the apobaraminic category can be useful especially in cases where the included holobaramins possess specimens with considerable similarity across holobaraminic boundaries. So humans can be compared with the organisms structurally and functionally most similar to them, namely chimps and gorillas.
Polybaramins
The fourth term, polybaramin (poly, from Greek for many), is employed for another mixture of unrelated organisms. It has been defined as a group (two or more specimens) consisting of part of at least two holobaramins. It may be any of numerous hodgepodges which could contain holobaramins, monobaramins, apobaramins, and individual specimens (see Figure 8).
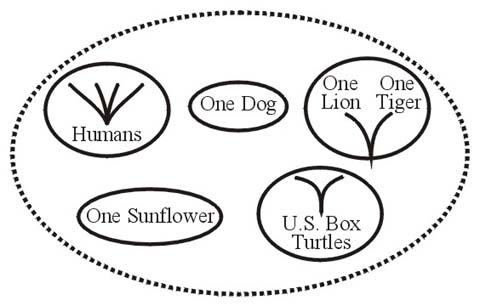
Figure 8.
As an example, a polybaramin could contain representatives of all human races, the two species of United States box turtles, one dog, one lion, one tiger, and one sunflower plant. The humans constitute a holobaramin. The box turtles belong to a monobaramin in a turtle holobaramin, the dog to a different monobaramin (in the canid holobaramin), the lion and tiger to another monobaramin (in the felid holobaramin), and the sunflower to a monobaramin within a plant holobaramin.
The adjective polybaraminic refers to the association between or among some or all parts of a polybaramin. For example, the sunflower and the human holobaramin would be polybaraminic, as would be the sunflower when compared with a box turtle monobaramin and a dog.
If an investigator is dealing with a polybaramin his taxonomic goal should be to separate its parts into the other three categories as soon as possible. Each of the monobaramins would need to be subtracted from this polybaramin and then separately studied carefully in order to determine their place in their respective holobaramins. The dog and the sunflower each is part of its own different holobaramin, and each of these holobaramins is unrelated to any other holobaramin in this polybaramin. Also, the humans should be analyzed individually to ascertain their proper relations within a holobaramin.
Further Thoughts
To repeat and expand this somewhat further, the Darwinian macroevolution model is represented by a single tree of relationships, every form of life being related to every other form of life (Figure 1). In the baraminic model there is a forest of trees without connecting roots (Figure 2). One of these rootless trees would have branches representing only human diversification, another for canids, another for felids, etc.
For people reared on an evolutionary diet, the above menu can be difficult to swallow and digest, because students of biology have been taught to think genetic relationship rather than genetic discontinuity. But there is a lack of evidence for connecting any holobaraminic group to any other holobaraminic group. This is true for both extinct and extant types of life.
It is common for scientists to utilize trees to depict relationships, but baraminologist David Cavanaugh believes that “trees” possibly may not be the best ways to portray relationships, but "other structures, such as networks or lattices may do a better job within many holobaramins. Tools of pattern recognition, such as projection plots, may perhaps be even better methods" (Cavanaugh, 1999-2000). So it remains to be seen just how relationships popularly will be represented in the future.
Baraminic Terminology
The four terms, holobaramin, monobaramin, apobaramin, and polybaramin formally and publicly were introduced by Walter ReMine (1990) at the Second International Conference on Creationism in Pittsburgh, PA 30 July 1990. Later in the week of the same conference Kurt Wise (1990), who had had extensive interchange with ReMine since 1983, endorsed ReMine’s discontinuity systematics, wedded it to his own young-Earth creation position, and stated that the name of this new systematic procedure was “baraminology.”
Wise also introduced a fifth term, namely archaebaramin, which could be conceived as the originally-created individual(s) of each holobaramin. For humans, Adam and Eve would constitute the archaebaramin. Two other terms which Wise has introduced are neobaramin which refers to living forms of life and paleobaramin for the older organisms. Archaebaramins are the most theoretical (Wise, 1990); whereas holo-, mono-, apo-, poly-, paleo- and neobaramins are to be determined on the basis of observational information.
Baraminology in Action
It is important to emphasize that the strictly empirical component of baraminology is discontinuity systematics which can be utilized by itself without any reference to religious literature. In fact, most of the sections in this present article, including the figures, actually are based on discontinuities as observed in nature. ReMine (2000) has pointed out that discontinuity systematics is intentionally designed to be a neutral, scientific method for studying some of nature’s patterns. We do not begin by assuming discontinuity; rather we follow the data to identify the discontinuities, wherever they may be. This systematic method is an empirical, scientific enterprise--moved by the data, not by theoretical presuppositions.
In the actual process of moving toward the goal of characterizing holobaramins, the taxonomist needs to identify apobaramins and partition them. Subtractive criteria need to be used in dividing the apobaramins into separate holobaramins. Then with the goal of characterizing holobaramins, the taxonomist focuses on the monobaramins, and additive criteria are employed to build these monobaramins.
An analogy for explaining this process has been proposed:
It is like there has been a huge snowfall covering the trees to the top, and we are digging down into the snow to identify the connections, the branches, limbs, and trunk. Is there one tree below? Or is it an orchard of separate distinct trees? As the data slowly come into view we will have arguments about what is connected to what, or whether there is discontinuity at a given place. Some researchers will mis-identify various branches as connected, when these later are seen as unconnected, and so forth. But this clears up as we dig. We are not “cutting and pruning” the data. Rather, we leave the data precisely where it is. We merely are cutting and pruning our perceptions--particularly our temporarily mistaken perceptions of the data (ReMine, 2000).
In other words the scientist is iterating tentative taxonomies by increasing or decreasing sizes of the branches to arrive at the best approximation of reality. This systematic procedure is driven by observed facts rather than some presupposed framework.
The goal of baraminology is to characterize holobaramins, but baraminologists do not recognize holobaramins as absolutely distinct from either apobaramins or monobaramins. Apobaramins contain one or more holobaramins. So if an apobaramin has been partitioned and only one holobaramin remains is that holobaramin still an apobaramin? The baraminologist says yes.
Also, if there is a portion (branch) of a holobaramin it is termed a monobaramin. This monobaramin will grow in size and complexity as more specimen branches are added. When a taxonomist has added all the branches which can be found among currently-living or extinct organisms, the taxonomist may judge the tree to be complete according to all currently-existing and applicable information. This means that the “tree”, which could have one or more branches, has all the forms of life believed to share genetic relationship, that is to say that are related by descent. So this group now by definition would be the holobaramin. Is it still a monobaramin? The baraminologist says yes! So how can a group be a monobaramin or an apobaramin and be a holobaramin at the same time?
Remine (1993, p. 447) and Wise (1999-2000) explain that we can think in terms of set theory. Consider a large circle, A (apobaramin). An inner portion of it is a smaller circle B (holobaramin), and this includes a still smaller inside region C (monobaramin). See Figure 9. Both the apobaramin (A) and monobaramin (C) are being changed in the direction of the middle circle, holobaramin (B).
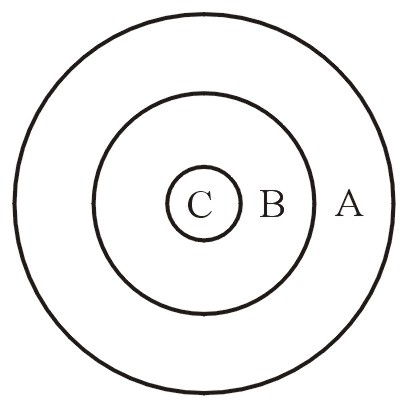
Figure 9.
Guidelines
In accomplishing the goal of separating parts of polybaramins, partitioning apobaramins, building monobaramins and characterizing holobaramins, a taxonomist needs guidelines for deciding what belongs to a particular monobaraminic branch. These standards will vary depending upon the groups being considered, but general guidelines which have been utilized include:
Scripture claims (used in baraminology but not in discontinuity systematics). This has priority over all other considerations. For example humans are a separate holobaramin because they separately were created (Genesis 1 and 2). However, even as explained by Wise in his 1990 oral presentation, there is not much relevant taxonomic information in the Bible. Also, ReMine’s discontinuity systematics, because it is a neutral scientific enterprise, does not include the Bible as a source of taxonomic information.
Hybridization. Historically Marsh and others have placed this criterion second only to the Bible; for if viable offspring could be obtained from a cross between two different forms, this would be definitive of their monobaraminic status. However, we realize today that the lack of known hybridization between two members from different populations of organisms does not necessarily by itself mean that they are unrelated. The hybridization criterion probably will retain validity, but it is being reconsidered in the light of modern genetics.
-
Ontogeny, namely the development of an individual from embryo to adult. Hartwig-Scherer (1998) suggested that comparative ontogeny followed hybridization in importance as a criterion for membership in a particular type.

Lineage. Is there evidence of a clear-cut lineage between and among either or both fossil and living forms.
Structure (morphology) and physiology (function). Structures may be macroscopic (large entities such as body organs), microscopic (small, and observed using magnification), and molecular (chemical) configurations.
Fossils in rock layers. These studies can include locations of fossil forms in the rock layers, and may entail considerations of Flood sediments.
Ecology. It is important to comprehend an organism’s niche, that is to say the region where it lives and how it interacts with the environment including other living things.
In order to determine baraminic distances among types of organisms it is important to utilize the most significant data. For instance, molecular studies with mitochondrial DNA and RNA were useful with some turtles, but the author questioned the baraminic utility of ecologic criterions (Robinson, 1997). In a baraminic study of human with non-human primates, the morphological (form) features such as teeth and bones as well as ecological characters including feeding and habitats were more valuable than chromosomal or molecular (hemoglobin and RNA) information (Robinson and Cavanaugh, 1998a). Also see Garcia-Pozuelo-Ramos, 1997; 1998; 1999. However, baraminic research on a broad spectrum of felids has revealed that ecological data were least reliable, and chromosomal data of low reliability, The morphological and molecular (protein and RNA) information were most important (Robinson and Cavanaugh, 1998b). For ongoing studies Cavanaugh (1999-2000) recently has emphasized that:
In particular, proteins, and their DNA patterns, which are part of fundamental cellular processes, have wide applicability in baraminological research. For example, proteins associated with cellular respiration like cytochrome C and cytochrome B are excellent candidates. See for example, the cytochrome C table in Denton (1986).
Statistics involving computer calculations are vital in the above investigations. It is to be expected that when baraminology is accepted widely, the science of taxonomy will be revolutionized. When systematists are dealing with a “forest” of trees rather than one large evolutionary tree it is possible that the categories of kingdom, phylum, division, class and even orders will be less useful in classification. However, among living things, groups of features within other groups of characteristics can be observed. These so called “nested patterns” (see ReMine, 1993; Wise, 1998) can extend beyond baraminic categories; so phenetic and cladistic methods may continue to be useful along with discontinuity systematics.
For those who have been steeped in Linnaean taxonomy and evolutionary thinking, discontinuity systematics may appear to be a preposterous proposal. However, this admittedly bold scheme should not be thought of as a departure from reality. Interestingly, on the first of August during the 1999 International Botanical Congress in St. Louis an overflow crowd heard a presentation promoting a so-called PhyloCode, a systematic scheme which would lead to the abolition of kingdoms, phylums, classes, orders, etc. (Milius, 1999). Also see papers by de Queiroz (1992; 1997a; 1997b). The proposed uncomplicated systematic procedure focuses on clades, each clade consisting of a single species and descendants of that species. In other words, the clade would be a holophyletic (genetically- united) group.
However, those utilizing a scheme such as this generally are thinking of clades within clades within larger clades on a macroevolutionary scale; whereas baraminology is more microevolutionary (small changes) which is much less speculative. For other taxonomic literature supporting typology see Scherer, 1993.
How many holobaramins will there be--3,000, 5,000, 10,000, 15,000, or more? At this time the best very tentative answer is, “probably in the low thousands”.
Active Baraminologists
On 6 March 1996 graduate student sent an email message to Kurt Wise suggesting a “baraminology study group”. The following day he emailed two other graduate students, Todd Wood and D. Ashley Robinson about this. The responses all were positive, and by 26 March 1996 the group had added Paul Nelson and John Meyer making a total of six. These men worked at identifying pertinent literature and establishing guidelines for the future. In June 1997, this group, plus more recent contacts David Cavanaugh and David Fouts, met in Dayton, TN where Wise worked, and together they established the Baraminology Study Group (BSG).
In addition to the above-mentioned eight men, there were Pete Williams, Jerry Kreps, and Jeff Tompkins who became involved to varying degrees. In October 1998 Joe Dasso and I accepted invitations to join the group, thus increasing membership to 13.
A three-day conference with the auspicious title "Baraminology ‘99: Creation Biology for the 21st Century" was organized and presented by the BSG in cooperation with the Departments of Biology and Chemistry at Liberty University in Lynchburg, VA, 5-7 August 1999. There were 24 invited participants.
Formal presentations were made by Todd C. Wood, D. Ashley Robinson, Kurt P. Wise, Pete J. Williams, and Paul Nelson. Topics related to baraminology included creation biology, phylogenetic inference, Biblical studies, design theory, the hybridization criterion, evolutionary webs, and non-systematics fields. The final afternoon was devoted to a workshop on baraminology of the family containing camels. These presentations and discussions on camels included an introduction, Biblical and linguistic studies, fossil record, hybridization, molecular studies, phylogenetic distortion, and conclusions. The final evening of the conference included a moderated discussion of “The Future of the BSG and Baraminology.”
Consideration was given to the establishment of a new society and a new journal. Participants volunteered for responsibilities centering on a second conference on the west coast in early 2001.
In spite of some differences of opinion, it was my impression that the group as a whole was very excited about progress made so far and prospects for the future of baraminology. Some of the scientists preferred to believe in an old Earth (billions of years), but the leadership and most of the attendees were united in maintaining a young Earth (thousands of years old) perspective. An introduction to baraminology and report on the 1999 conference has been written by an attendee, botanist Margaret Helder, 1999.
Concluding Comments
Baraminology may be thought of as a typological approach to classifying forms of life, both living and fossilized. In former centuries scientists theorized typologically more commonly than they do at the present time. However, because of the many difficulties (for example, convergences and reversals) which plague the macroevolutionary thinker, there is a growing receptivity to typology.
Baraminologists believe that they are at the forefront of modern progressive thinking. Those interested in learning more about the ten-year-old field of baraminology should consult the references.
Acknowledgments
David Cavanaugh, Kurt Wise and Walter ReMine read earlier drafts of this manuscript and contributed much invaluable information. Other aid came from John W. Cuozzo, Scott A. Mahathey, Todd Wood, and D. Ashley Robinson. Help with mechanical details has been rendered by Lane P. Lester, CRSQ Managing Editor. The paper was written in response to patient encouragement from the CRSQ Editor, Emmett L. Williams.
References
CRSQ: Creation Research Society Quarterly
Bartz, Paul A. 1991. A refinement of biosystematics which reflects baraminic variation. CRSQ 28(1):18-20.
Cuozzo, Jack. 1998. Buried alive: the startling truth about Neanderthal Man. Master Books, PO Box 727, Green Forest, AR 72638.
___. 2000. Personal communications.
Denton, Michael. 1986. Evolution: a theory in crisis. Woodbine House, Rockville, MD.
de Queiroz, Kevin. 1992. Phylogenetic taxonomy. Annual Review of Ecology and Systematics 23:449-480.
___. 1997a. Misunderstandings about the phylogenetic approach to biological nomenclature: a reply to Liden and Oxelman. Zoologica Scripta 26(1):67-70.
___. 1997b. The Linnaean hierarchy and the evolutionization of taxonomy, with emphasis on the problem of nomenclature. Aliso (The Rancho Santa Ana Botanic Garden, Claremont, CA 91711-3157) 15(2):125-144.
Frair, Wayne. 1982. Serum electrophoresis and sea turtle classification. Comparative Biochemistry and Physiology 72B:1-4.
___. 1991. Original kinds and turtle phylogeny. CRSQ 28(1):21- 24.
___. 1999. Creationist classification--an update. Creation Matters 4(1):1,5.
Garcia-Pozuelo-Ramos, Celedonio. 1997. Taxonomy of primates. CRSQ 34(2):93-102.
___. 1998. Dental variability in the domestic dog (Canis familiaris): implications for the variability of primates. CRSQ 35(2):66-75.
___. 1999. Craniodental variability in the domestic dog (Canis familiaris) and its implications for the variability in primates. CRSQ 36(3):116-123.
Hartwig-Scherer, Sigrid. 1998. Apes or ancestors?: interpretations of the hominid fossil record within evolutionary and basic type biology. In William A. Dembski, editor. Mere creation: science, faith and intelligent design, pp. 212-235. Intervarsity Press, Downers Grove, IL.
Helder, Margaret. 1999. Introducing baramins. Creation Dialogue (5328 Calgary Trail South, Suite 1136, Edmonton, Alberta T6H 4J8) 26(3):6-7.
Hirayama, Ren. 1998. Oldest known sea turtle. Nature 392:705-708.
Iverson, John B. 1992. A revised checklist with distribution maps of the turtles of the world. Privately printed, Richmond, IN.
Klein, Richard G. 1999. The human career : human biological and cultural origins. The University of Chicago Press, Chicago, IL.
Lutz, Peter L., and John A. Musick. 1997. The biology of sea turtles. CRC Press, New York.
Marsh, Frank L. 1941. Fundamental biology. Published by the author, Lincoln, NE.
___. 1969. The form and structure of living things. CRSQ 6(1):13-25.
Milius, Susan. 1999. Should we junk Linnaeus? Science News 156(17):268-270.
O’Brien, Stephen J. 1997 (June). The human-cat connection. National Geographic 191(6):77-85.
ReMine, Walter J. 1990. Discontinuity systematics: a new methodology of biosystematics relevant to the creation model. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 207-216. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1993. The biotic message: evolution versus message theory. St. Paul Science, PO Box 28006, Saint Paul, MN 55128. [up]
___. 2000. Personal communications.
Robinson, D. Ashley. 1997. A mitochondrial DNA analysis of the testudine apobaramin. CRSQ 33(4):262-272.
Robinson, D. Ashley, and David P. Cavanaugh. 1998a. A quantitative approach to baraminology with examples from the catarrhine primates. CRSQ 34(4):196-208.
___. 1998b. Evidence for a holobaraminic origin of the cats. CRSQ 35(1):2-14.
Scherer, Siegfried. 1993. Typen des Lebens. Pascal Verlag, Berlin.
Williams, Pete J. 1997. What does min mean? Creation Ex Nihilo Technical Journal 11(3):344-352.
Wise, Kurt P. 1990. Baraminology: a young-Earth creation biosystematic method. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345-360. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1992. Practical baraminology. Creation Ex Nihilo Technical Journal 6(2):122-137.
___. 1998. Is life singularly nested or not? In R. E. Walsh, editor, Proceedings of the Fourth International Conference on Creationism. Technical Symposium Sessions, pp. 619-631. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1999-2000. Personal communications.
Reprints Available
Reprints of “Baraminology—Classification of Created Organisms” are available as a separate publication . Individual copies are $2.00 each. Add 15% for postage (minimum $3.00, maximum $10) Mail your order to: Creation Research Society, P.O. Box 8263, St. Joseph, MO 64508-8263.
Author: Wayne Frair, Ph.D. Text published in Creation Research Society Quarterly, Vol. 37, No. 2 (September 2000), pp. 82-91. Provided by Creation Research Society.
Copyright © 2001, Creation Research Society, All Rights Reserved—except as noted on attached “Usage and Copyright” page that grants ChristianAnswers.Net users generous rights for putting this page to work in their homes, personal witnessing, churches and schools. All photos were provided by Films for Christ and are copyrighted.