Что имеется в виду в книге Бытие под «родом»? Бараминология - классификация сотворённых организмов
Смотрите эту страницу в: Английский (English)

Десятилетиями креационисты употребляли слова род», «тип» или «группа» для того, что они представляли как категории генетически неродственных организмов, включая те, что были созданы Творцом в течение Недели Сотворения. Предполагалось, что внутри каждой из этих категорий от общих первопредков произошли разнообразные виды, подвиды и породы. Однако, до последних лет не было серьёзной всеобъемлющей методики классификации, направленной на характеристику каждой сходной категории, которая отделена от всех остальных категорий генетическим разрывом. Теперь развитие бараминологии (с прерывистой систематизацией) сделало её плодотворным подходом к классификации в рамках модели сотворения. Разработаны её терминология и методология, а летом 1999 г. была проведена первая бараминологическая конференция.
[От редактора: Термин бараминология означает изучение сотворённых родов. Он создан из древнееврейских слов «бара» (сотворил) и «мин» (род).]
Введение
Среди основных механизмов человеческого мышления находятся классификация и отождествление. Люди занимаются ими постоянно, и они существенно важны для нашей жизни. Люди научились классифицировать мысли и вещи, и в процессе своего взаимодействия с окружающим миром они непрестанно отождествляют данные органов чувств, такие как запахи, зрительные впечатления (небо, пища, животное, растение и т.д.), звуки (вой сирены, лай собаки, музыка и т.д.), вкус (сладкий, кислый, и т.д.) и другие ощущения, напр., шероховатость и гладкость. Сложность всего этого поразительна.
Интересно то, что Библия рассказывает о том, как Бог привёл к Адаму всех наземных животных и птиц (Бытие 2:19), чтобы Адам посмотрел на них и назвал их. Эта история может иметь более глубокий смысл, чем тот, что открывается при поверхностном прочтении. В процессе наречения этих существ Адам узнал нечто о самом себе. Он осознал свою способность к восприятию и к распознаванию образцов. Некоторые создания были одеты в чешую, а некоторые в шерсть или в перья.
Кроме того, Адам должен был что-то узнать от Бога - Дизайнера. В творении он мог наблюдать выражение рукоделия Господа. Открыв для себя природу, Адам никогда не остался бы таким, как раньше. Сцена была теперь готова, и вслед за этим миллиарды студентов, изучающих естествознание, должны были сыграть свою роль на сцене жизни, и при этом ощутить восхищение от раскрытия в природе послания от Бога (Псалом 19:1; Римлянам 1:19,20; и см. ReMine, 1993).
Таксономия и систематизация
При размышлении обо всём этом нам станет легче осознать, насколько важна классификация в биологическом изучении ископаемых и существующих ныне форм жизни. Таксономия - это термин, которым обозначается наука, занимающаяся классификацией живых существ по их естественным группам. В основном сегодняшние учёные пользуются таксономической системой, введённой шведским ботаником Линнеем около 250 лет назад.
Однако, при изучении моделей жизни и классификации возможны различные подходы. Они называются таксономическими системами и объединяются под заголовком виды систематизации. Для изучения природных моделей Ри-Майн (1993, p. 444) сравнил различные процедуры с функциями разных типов плёнки (инфракрасного, ультрафиолетового и рентгеновского). Например, есть филетическая или эволюционная систематизация (или дарвиновская), в которой предполагается макроэволюция (предполагающая крупные изменения). См. Рисунок 1.
Систематические модели, которые менее акцентируют эволюцию, можно назвать фенетическими, потому что они основаны на внешних чертах организмов, а не обязательно на их возможных крупномасштабных эволюционных соотношениях. Существует также кладистический подход, в котором внимание концентрируется на общих характеристиках. Важнее всего то, что все популярные филетические, фенетические и кладистические методики были адаптированы к макроэволюционным рассуждениям.
Самыми ранними методами были фенетические, которые имели явное преимущество - они были основаны на объективных наблюдениях, и их можно было проще всего проверить. Критики базовой фенетики чувствуют, что её простота не способствует адекватному рассмотрению исторических причин и следствий. Тем не менее, те, кто пользуются бараминологией (с разрывной систематизацией), как представлено в этой статье, могут и не знать популярных фенетической и кладистической методик, но, по крайней мере, они употребляют некоторые фенетические методы как одно из своих орудий.
Разрывная систематизация и бараминология
Учёные, предпочитающие что-то отличное от макроэволюционной модели, недавно разработали то, что, как они считают, является более реалистической систематизацией, основанной на разрывах или типологию, обнаруживаемую в природе. Эта методика была соответственно названа разрывной систематизацией (официально представлена Вальтером Ри-Майном Walter J. ReMine, 1990), или, в сочетании с библейским откровением, бараминологией (термин введён Куртом П. Визе Kurt P. Wise, 1990). Бараминологию можно определить как таксономию на основе сотворённых родов (см. Bartz, 1991; Frair, 1991; 1999; и Рисунок 2).


Слово «барамин» было придумано Фрэнком Л. Маршем и впервые опубликовано в 1941 г.; оно произведено от древнееврейского глагола бара (творить) и слова мин (род) (также см. Marsh, 1969; Williams, 1997).
Поскольку классификация лежит в основе всех биологических исследований, довольно значимо то, что креационисты теперь уделяют этому вопросу очень большое внимание. С 1990 г. были достигнуты серьёзные успехи, и бараминологи разработали собственную терминологию, которая в настоящее время представляется вполне пригодной для систематических исследований. Главная цель бараминологии - в том, чтобы определить, какие организмы имеют общее происхождение.
Марш использовал термин барамин в общем смысле для обозначения целой группы известных, неизвестных и, возможно, подразумеваемых организмов, объединённой общими генетическими отношениями. Но теперь он конкретизирован, и в четыре нынешние основные бараминные группы могут быть включены только те организмы, которые доступны изучению в живом или ископаемом видах (включая окаменелости). Для четырёх наиболее важных бараминных категорий используются термины холобарамин, монобарамин, апобарамин и полибарамин.
Холобарамины
Основным термином бараминологии является холобарамин (от греческого холос - целый). Холобарамин - это все или только известные существующие ныне и/или вымершие формы жизни, которые понимаются как имеющие общие генетические отношения. Это целостная группа, которая считается объединённой общим родством.
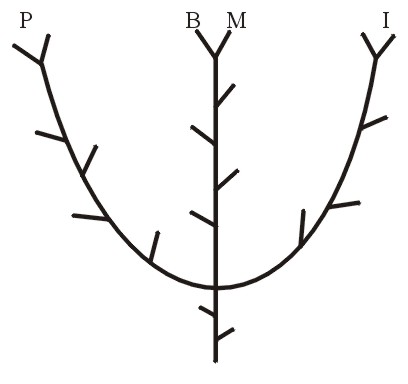
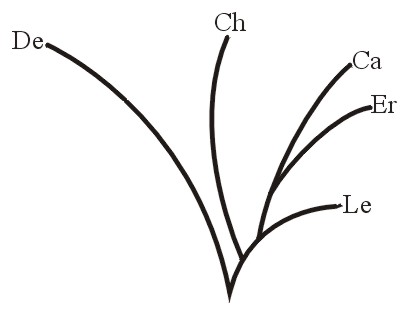
Поэтому теперь каждая естественная группа родственных растений или родственных животных составляет холобарамин; или, если использовать более конкретную креационистскую терминологию, холобарамин состоит из всех известных организмов в группе, начиная с сотворённых Богом первоначальных организмов (см. Wise, 1992). Холобарамин может быть представлен как ветвящееся дерево, причём узлы и кончики веток представляют собой всех известных членов (подвиды, виды и т.д.) «рода» («группы» или «типа»). См. Рисунок 2. При сравнении отдельных представителей или групп очевидно родственных организмов их можно обозначить как холобараминные, если они являются частями одного холобарамина.
За несколько последних десятилетий исследователи-креационисты обычно употребляли термины «род», «группа» или «тип» взаимозаменяемо; или же отдельные исследователи предпочитали то или иное конкретное общее слово для «бараминов» Марша, а также для того, что можно сейчас обозначить как холобарамины. Теперь при систематизации конкретных таксонов растений или животных можно отбросить более старую терминологию и построить их деревья, показывающие холобараминные родовые сходства, и таким образом, границы общего происхождения.
Важным примером холобарамина могут служить люди, Homo sapiens. На концах холобараминных ветвей находятся различные расы (кавказцы, эфиопы, монголы, американские индейцы и т.д.). См. Рисунок 3. Член любой из этих рас потенциально может иметь общее потомство с супругом противоположного пола из любой другой расы.
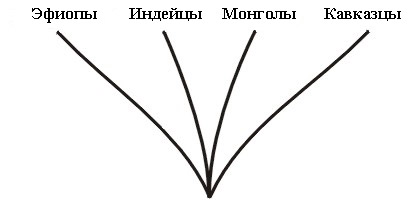
Часто в антропологической литературе можно встретить упоминание о более чем восьми человеческих географических расах, даже с дополнительными промежуточными популяциями. Однако, в настоящей статье я намерен не вдаваться в дискуссии о различных возможностях того, что выражено здесь в тексте или на каком-либо из рисунков, а просто проиллюстрировать использованные таксономические принципы.
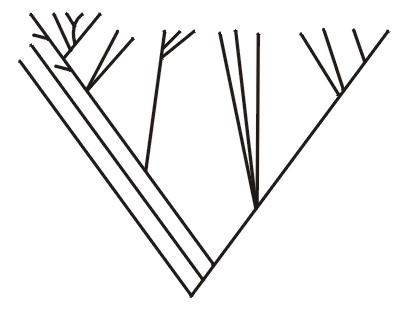
Ещё один холобарамин может состоять из морских черепах (см. Wise, 1992; Robinson, 1997). Схему, изображающую общие формы живых и вымерших морских черепах, можно найти в Lutz and Musick, 1997, p. 8. Такая схема называется «кладограммой» и основана на исследованиях специалистов Гэффни и Мейлана, но не все авторитеты согласны с их оценкой имеющихся данных. См. также Hirayama, 1998. Рисунок 4 представляет собой очень обобщённое изображение всех живых и вымерших морских черепах.
Во всех этих типах исследований настоящая цель разрывной систематизации состоит в том, чтобы с помощью эмпирических фактов определить границы общего родства и, таким образом, сойтись по вопросу о холобараминах.
Различные члены холобарамина могли произойти в результате передачи потомству различных генов (ДНК) от родительских организмов. Это часто происходит сегодня. Или же, со времени сотворения могли произойти какие-то наследственные видоизменения ДНК (мутации), а они были переданы разным ветвям потомства. Естественный отбор мог повлиять на потенциал выживаемости различных потомков одних родителей.
Монобарамины
Второй термин, которым пользуется бараминология, - это монобарамин (от греческого моно - один или единственный). Термин монобарамин определяется Ри-Майном (1993, p. 444) как:
группа, состоящая только из организмов, объединённых общим происхождением, но не обязательно всех из них. (Группа, включающая в себя один целый холобарамин или его часть).
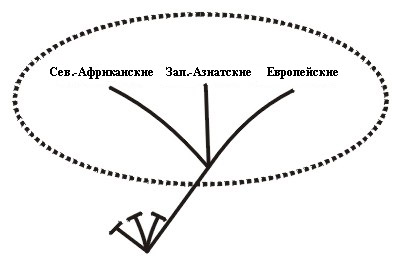
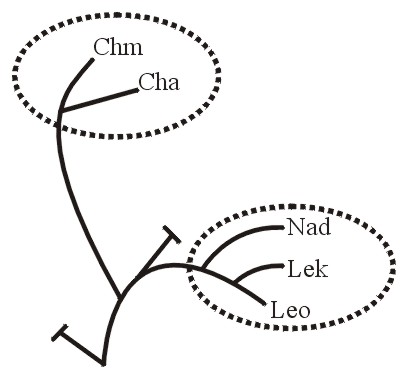
Если холобарамин представить как дерево, одна или несколько из его ветвей будут монобараминами. Например, среди людей кавказцы будут монобарамином (Рисунок 5).
Или же, в случае с морскими черепахами, пять ныне существующих типов, обитающих в океанах по всему миру, составляют монобарамин (Рисунок 6A из Frair, 1982; см. тж.Iverson, 1992, p. 80). Также и группа зелёных черепах, Chelonia, или ветвь, в которую входит черепаха Ридли, Lepidochelys, - каждая из них будет монобарамином (Рисунок 6B). Отдельные представители или группы могут считаться монобараминными, если они представляют собой части одного холобарамина (Рисунки 5, 6а, 6б, and 7б).
Кроме того, проводилось систематическое изучение конкретных монобараминов, в которых возникли различные формальные признаки (такие как внешность наиболее узнаваемых видов, подвидов, пород и т.д.). Исследования монобараминов в основных чертах не различаются в зависимости от философии систематизации, которой придерживаются учёные. Различие между (1) филетической точкой зрения (в смысле дарвиновской макроэволюционной перспективы) и (2) точкой зрения бараминной разрывной систематизации (креационистской микроэволюционной концепции «ограниченных изменений») будет, главным образом, в том, что первая из них пользуется эмпирическими данными для того, чтобы экстраполировать их на каких-то воображаемых более ранних предков. Но бараминолог утверждает, что представление о филогенезе не должно выходить за пределы убедительных фактов, и что учёных надо освободить от чувства обязательства философски конструировать обширные схемы филогенеза (эволюционные деревья) при отсутствии неопровержимых фактов.
Апобарамины
Третьим бараминологическим термином является апобарамин (от греческого апо - от, прочь), и им обозначается «группа, состоящая из не менее, чем одного полного холобарамина» (Wise, 1999-2000). Она может включать всего один холобарамин или более одного холобарамина. «Но в ней должно быть не менее одного холобарамина полностью». Ни один организм, входящий в холобарамин в рамках одного апобарамина, не имеет общих предков ни с одним организмом за пределами своего холобарамина (это основано на определении, данном холобарамину). См. Рисунок 7а.
Прилагательное апобараминный относится к отношениям между явно неродственными группами (холобараминами). Например, всё человечество как единая группа будет апобараминной, поскольку ни один из её членов не имеет общих предков ни с какими другими организмами. Группа из всех людей и всех черепах также будет апобараминной, потому что ни один человек и ни одна черепаха не имеет общих предков ни с какими организмами, не являющимися людьми или черепахами.

|
Считается, что все лошади (лошади, ослы и зебры) родственны, потому что они могут скрещиваться, а следовательно, принадлежат к одному холобарамину. Кроме того, есть «собачий» холобарамин с монобараминными ветвями волков, гиен, койотов, шакалов и сотен пород домашних собак. «Кошки» составляют ещё один холобарамин с мономараминными ветвями львов и тигров, пум, рысей, домашних кошек и т.д. (см. O’Brien, 1997). Группа из всех лошадей (семейство лошади), из всех собак (семейство собачьи) и всех кошек (семейство кошачьи) будет апобараминной, потому что ни одна лошадь, собака или кошка не имеют родственных отношений ни с какими организмами, которые не являются лошадью, собакой или кошкой.
 |
Черепаший апобарамин может состоять из одного, двух, трёх или четырёх холобараминов (см. Wise, 1992). В настоящей статье я рассматриваю морских черепах как составляющих холобарамин; следовательно, группа, в которую входит холобарамин морских черепах, все лошади, все собачьи и все кошачьи будет апобараминной, поскольку ни один из членов любого из этих четырёх холобараминов не имеет родственных отношений ни с какими представителями за пределами их соответственных холобараминов.
Термин апобарамин особенно полезен при рассмотрении двух типов организмов (попарные сравнения). Использование попарных сравнений - это наиболее обычная таксономическая процедура.
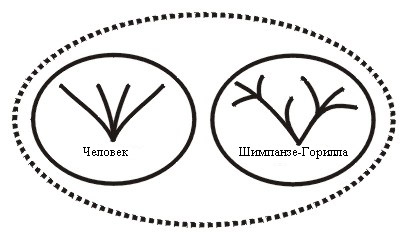
Например, нынешний Отряд приматы включает в себя человекообразных обезьян, обезьян, лемуров и людей. Все человеческие расы принадлежат к одному холобарамину; в то время как шимпанзе с гориллами являются членами другого холобарамина.
 |
Поэтому группа из человеческого холобарамина и холобарамина шимпанзе и горилл будет апобараминной (см. Рисунок 7а). Далее, объединение человеческого холобарамина с любым или всеми из других обезьяньих холобараминов будет апобараминным. Никто из членов этих холобараминов не будет иметь общих предков ни с одним членом любого другого холобарамина, входящего или даже не входящего в этот апобарамин. См. бараминное исследование приматов в Robinson and Cavanaugh, 1998a.
Выводы о холобараминных отношениях шимпанзе и горилл (Рисунки 7а и 7б) основаны на Hartwig-Scherer, 1998.
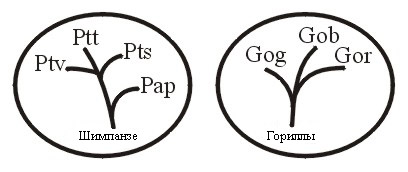
Кавано (1999-2000) также сообщил мне, что повторное изучение данных из работы Робинсона и Кавано (Robinson, Cavanaugh, 1998a) подтверждает холобараминный статус шимпанзе и горилл. Однако, как объяснил Кляйн Klein (1999, pp. 135-136), летопись окаменелостей даёт слишком мало полезной информации для понимания истории развития шимпанзе и горилл. Когда будет убедительно установлено, что окаменелости относятся к шипанзе и гориллам, их надо будет отнести к их холобарамину. Возможно, даже группа шимпанзе и горилл должна быть разделена на два отдельных холобарамина. Рисунок 7а показывает, как может использоваться апобараминная категория, особенно в тех случаях, когда входящие в неё холобарамины имеют в своём составе представителей со значительными сходствами по обе стороны холобараминных границ. Поэтому людей можно сравнить с организмами, по строению и функциям наиболее схожими с ними, а именно с шимпанзе и гориллами.
Полибарамины
Четвёртый термин, полибарамин (от греческого поли - много), употребляется для ещё одного сочетания неродственных организмов. Он определяется как группа (два или более представителя), состоящая из не менее, чем двух холобараминов. Здесь возможны самые разнообразные смеси, содержащие холобарамины, монобарамины, апобарамины и отдельных их представителей (см. Рисунок 8).
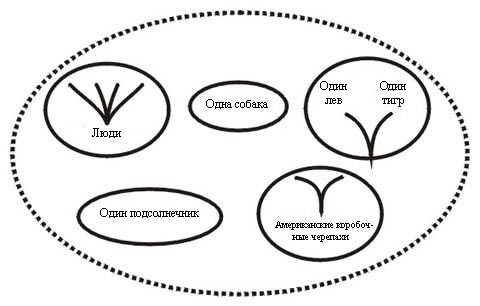
Например, в полибарамин могут входить представители всех человеческих рас, два вида американских коробочных черепах, одна собака, один лев, один тигр и один подсолнечник. Люди составляют холобарамин. Коробочные черепахи принадлежат к одному монобарамину в составе черепашьего холобарамина, собака - к другому монобарамину (в составе монобарамина собачьих), лев и тигр - ещё к одному монобарамину (в составе холобарамина кошачьих), а подсолнечник - к монобарамину в составе холобарамина растений.
Прилагательное полибараминный относится к связям между некоторыми или всеми частями полибарамина. Например, подсолнечник и человеческий холобарамин будут полибараминными, то же касается и подсолнечника по отношению к монобарамину коробочных черепах и собаке.
Если исследователь имеет дело с полибарамином, его таксономической целью должно быть как можно более скорое разделение его частей на три другие категории. Надо будет вычесть из этого полибарамина каждый из монобараминов, а потом тщатльно изучать их отдельно, чтобы определить их место в соответствующих холобараминах. И собака, и подсолнечник являются частью других холобараминов, и каждый из этих холобараминов неродственен любому другому холобарамину в этом полибарамине. Также и людей следует анализировать отдельно, чтобы выявить их собственные взаимооотношения внутри холобарамина.
Дальнейшие рассуждения
Чтобы повторить и немного далее расширить сказанное выше, вспомним, что дарвиновская модель макроэволюции представлена единым деревом взаимосвязей, причём каждая форма жизни родственна всем остальным формам жизни (Рисунок 1). В бараминной модели мы имеем лес из деревьев без соединяющих их корней (Рисунок 2). Одно из этих деревьев без корней будет иметь ветви, представляющие только разнообразие людей, другое - кошачьих, третье - собачьих и т.д.
Для людей, вскормленных на эволюционном рационе, изложенное выше меню может быть трудно проглотить и переварить, потому что студентов-биологов учили думать о генетических взаимосвязях, а не о генетических разрывах. Но имеется недостаток фактов, связывающих любую из холобараминных группы с любой другой холобараминной группой. Это верно и в отношении к нынешним типам жизни, и к вымершим.
Учёные часто используют деревья для того, чтобы изобразить взаимоотношения, но бараминолог Дэвид Кавано полагает, что «деревья» - это, возможно, не лучший способ показывать взаимосвязи, но «другие структуры, такие как сети или решётки, могут в случае с многими холобараминами пригодиться больше. Инструменты распознавания узора, такие как проектировочные чертежи, могут, возможно, оказаться даже лучшими методами» (Cavanaugh, 1999-2000). Поэтому нам остаётся только увидеть, как взаимоотношения будут популярно представляться в будущем.
Бараминная терминология
Уолтером Ри-Майном на Второй Международной конференции по креационизму в Питтсбурге (штат Пенсильвания) 30 июля 1990 г. были формально и публично введены четыре термина: холобарамин, монобарамин, апобарамини полибарамин. Позднее, в течение недели на этой конференции Курт Визе (1990), с 1983 г. поддерживавший интенсивные контакты с Ри-Майном, поддержал ри-майновскую разрывную систематизацию, вплёл её в свою собственную креационистскую теорию молодости Земли и объявил, что эта новый метод систематизации называется «бараминология».
Визе ввёл также пятый термин, а именно архебарамин, который можно понимать как первоначально сотворённого представителя или представителей каждого холобарамина. В случае с людьми архебарамин будут составлять Адам и Ева. Визе ввёл и два других термина: необарамин, который относится к живым формам жизни и палеобарамин для более древних организмов. Археобарамины носят наиболее умозрительный характер (Wise, 1990); в то время как холо-, моно-, апо-, поли-, палео- и необарамины должны определяться на основе данных наблюдения.
Бараминология в действии
Важно подчеркнуть, что строго эмпирический компонент бараминологии - это разрывная систематизация, которую можно применять и саму по себе, безотносительно ко всякой религиозной литературе. Действительно, большинство разделов данной статьи, включая цифры, в сущности, основаны на разрывности, наблюдаемой в природе. Ри-Майн (2000) указал, что разрывная систематизация разработана как нейтральный, научный метод для изучеия некоторых природных моделей. Мы не исходим из предположения о разрывности: мы, скорее, следуем данным для того, чтобы выявить разрывы, где бы они ни наблюдались. Этот метод систематизации является эмпирическим, научным предприятием на основе фактов, а не теоретических предположений.
В ходе самого процесса движения к цели характеристики холобараминов, таксономисту нужно выявить апобарамины и расчленить их. При делении апобараминов на отдельные холобарамины надо использовать критерии вычитания. Затем с целью характеристики холобараминов, таксономист концентрирует внимание на монобараминах, и для построения этих монобараминов применяются критерии дополнения.
Для объяснения этого процесса предлагалась такая аналогия:
Это похоже на то, как если бы прошёл очень сильный снегопад, покрывший деревья доверху, и мы раскапываем снег, чтобы выявить сочленения, ветви, сучья и ствол. Одно ли дерево там внизу? Или же целый сад отдельных деревьев? По мере того, как медленно становятся видны данные, у нас будут появляться доводы о том, что с чем связано, или же о том, что в данном месте есть разрывность. Некоторые исследователи неверно посчитают, что различные ветви связаны, когда в действительности они окажутся несвязанными, и так далее. Но это выясняется по мере раскопок. Мы не «подрезаем и постригаем» данные. Скорее, мы оставляем данные точно там, где они есть. Мы просто подрезаем и постригаем наши представления - особенно наше временно ошибочное восприятие фактов (ReMine, 2000).
Другими словами, учёный пробует всё новые и новые таксономии, увеличивая или уменьшая размеры ветвей, чтобы достичь наилучшего приближения к реальности. Этот систематический метод движется наблюдаемыми фактами, а не предполагаемой моделью.
Целью бараминологии является характеристика холобараминов, но бараминологи не считают, что холобарамины совершенно отличны от апобараминов или монобараминов. Апобарамины содержат в себе не менее одного холобарамина. Поэтому если апобарамин разделить, и от него останется только один холобарамин, то является ли этот холобарамин по-прежнему апобарамином? Бараминолог говорит «да».
Также, если имеется часть (ветвь) холобарамина, то она называется монобарамином. Этот монобарамин будет увеличиваться в размере и в сложности по мере добавления новых ветвей. Когда таксономист добавил все ветви, которые можно найти среди ныне живущих или вымерших организмов, таксономист может считать дерево полным, согласно всей имеющейся ныне и доступной информации. Это означает, что на «дереве», на котором может быть одна или больше ветвей, имеются все формы жизни, которые считаются генетическими родственными, то есть, имеющими общее происхождение. Поэтому теперь эта группа по определению будет холобарамином. Монобарамин ли это? Бараминолог говорит «да». Так как же одна и та же группа может быть и монобарамином или апобарамином, и одновременно холобарамином?
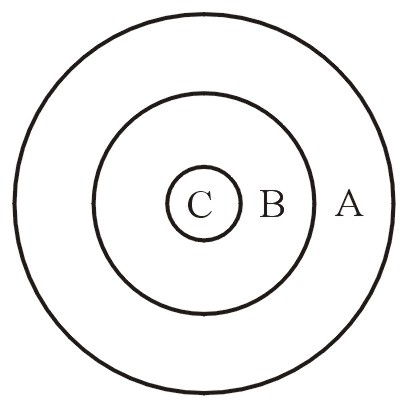
Ри-Майн (1993, p. 447) и Визе (1999-2000) объясняют, как здесь можно рассуждать в рамках изложенной теории. Представьте себе большой круг, А (апобарамин). Внутренняя часть его представляет собой меньший круг В (холобарамин), а в нём есть ещё меньший участок С (монобарамин) См. Рисунок 9. И апобарамин (A), и монобарамин (С) изменяются в направлении среднего круга, холобарамина (В).
Критерии
Преследуя цели выделения частей полибараминов, разделения апобараминов, построения монобараминов и характеристики холобараминов, таксономист нуждается в критериях для определения того, что относится к конкретной монобараминной ветви. Эти стандарты будут варьироваться в зависимости от рассматриваеиых групп, но общие использовавшиеся критерии включают в себя:
Утверждения Писания (используются в бараминологии, но не в разрывной систематизации). Это соображение имеет приоритет над всеми другими. Например, люди - это отдельный холобарамин, потому что они были созданы отдельно (Бытие 1 и 2). Однако, даже при том, что объяснил Визе в своём устном докладе в 1990 г., в Библии не очень много таксономической информации по этому вопросу. Также и разрывная систематизация Ри-Майна, будучи нейтральным научным методом, не включает в себя Библию как источник таксономической информации.
Гибридизация. Исторически Марш и другие учёные ставили этот критерий только на второе место после Библии; ибо если от скрещивания между двумя различными формами можно получить жизнеспособное потомство, то это будет определять их монобараминный статус. Однако сегодня мы понимаем, что отсутствие известной гибридизации между двумя членами разных популяций организмов само по себе вовсе не обязательно означает, что они неродственны. Критерий гибридизации, вероятно, сохранится, но он пересматривается в свете современной генетики.
Онтогенез, а именно, индивидуальное развитие от эмбриона до взрослой особи. Хартвиг-Шерер (1998) предположил, что сравнительный онтогенез следует по степени важности за гибридизацией в качестве критерия для членства в определённом типе.

Родословная. Имеются ли свидетельства явного линейного происхождения друг от друга ископаемых или живых форм.
Строение (морфология) и физиология (функция). Строение может быть макроскопическим (крупные целостные части, такие как органы тела), микроскопическими (мелкими, наблюдаемыми с помощью увеличения), и молекулярными (химическими) конфигурациями.
Окаменелости в слоях породы. В эти исследования может входить расположение окаменелых форм в слоях породы, а вслед за ним - рассмотрение отложений, образовавшихся при Потопе.
Экология. Важно понять нишу, которую занимает организм, то есть регион его обитания и то, как он взаимодействует с окружающей средой, включая других живых существ.
Чтобы определить бараминные расстояния между типами организмов, важно использовать наиболее значимые данные. Например, молекулярные исследования митохондриальных ДНК и РНК были полезны в случае с некоторыми черепахами, но автор сомневается в бараминной полезности экологических критериев. (Robinson, 1997). При бараминном изучении человека и иных приматов, морфологические (формальные) черты, такие как зубы и кости, как и экологические характеристики, включая питание и среду обитания, были более ценными, чем хромосомные или молекулярные (гемоглобин и РНК) (Robinson and Cavanaugh, 1998a). См. также Garcia-Pozuelo-Ramos, 1997; 1998; 1999. Однако, бараминное исследование широкого спектра кошачьих выявило то, что экологические данные были самыми ненадёжными, а на хромосомные данные также можно было мало полагаться. Наиболее важной была морфологическая и молекулярная (белок и РНК) информация (Robinson and Cavanaugh, 1998b). Что касается нынешних исследований, то Кавано (1999-2000) недавно подчеркнул, что:
В частности, при бараминологическом исследовании широко применимы белки и их модели ДНК, которые участвуют в основных клеточных процессах. Например, отличными кандидатами были белки, связанные с клеточным дыханием, такие как цитохром С и цитохром В. См., напр., таблицу цитохрома С в Denton (1986).
В вышеупомянутых исследованиях крайне важна статистика с использованием компьютерных вычислений. Ожидается, что когда бараминология будет принята широко, в таксономической науке произодёт переворот. Когда систематизаторы будут иметь дело с «лесом» деревьев, а не с одним эволюционным деревом, возможно, категории царств, типов, классов и даже отрядов, будут менее полезны в классификации. Однако, среди живых существ можно наблюдать группы признаков в рамках других групп характеристик. Эти так называемые «гнездовые модели» (см. ReMine, 1993; Wise, 1998) могут выходить за пределы бараминных категорий; поэтому фенетические и кладистические методы могут и далее быть пригодными наряду с разрывной систематизацией.
Для тех, кто привык к линнеевской таксономии и эволюционному мышлению, разрывная систематизация может показаться нелепым предложением. Однако эта несомненно смелая схема не должна восприниматься как отход от реальности. Интересно, что 1 августа 1999 г. в ходе Международного конгресса ботаников в Сент-Луисе огромная толпа собралась на доклад о так называемом филокоде, схеме систематизации, которая привела бы к отмене царств, типов, классов, отрядов и т.д. (Milius, 1999). См. тж. работы де Кейроса: de Queiroz (1992; 1997a; 1997b). Предложенный несложный метод систематизации обращает основное внимание на «клады», причём каждый клад состоит из одного вида и потомства этого вида. Другими словами, «клад» будет холофилетической (генетически единой) группой.
Однако, те, кто пользуется такой схемой, обычно думают о «кладах» внутри «кладов» внутри больших «кладов» в макроэволюционном масштабе; в то время как бараминология более микроэволюционна (небольшие изменения), а значит, намного менее умозрительна. Другую таксономическую литературу, поддерживающую типологию, см. в Scherer, 1993.
Сколько будет холобараминов - 3 000, 5 000, 10 000, 15 000, или больше? В настояшее время лучше всего ответить будет так: «наверное, где-то несколько тысяч».
Активные бараминологи
6 марта 1996 г. студент выпускного курса университета Нил А. Доран отправил Курту Визе электронное письмо, предлагая образовать «группу по изучению бараминологии». На следующий день он написал об этом двум другим выпускникам - Тодду Вуду и Д. Эшли Робинсону. Все ответы были положительными, а 26 марта 1996 г. к группе присоединились Пол Нельсон и Джон Майер, в результате чего группа расширилась до шести человек. Эти люди работали над выявлением литературы, связанной с данным вопросом, и установлением критериев на будушее. В июне 1997 г. Доран, Визе, Вуд и Робинсон плюс их новые знакомые Дэвид Кавано и Дэвид Фаутс встретились в Дейтоне (штат Теннесси), где работал Визе, и вместе учредили Группу по изучению бараминологии (ГИБ).
Кроме вышеупомянутых восьми человек, в ней были Пит Вильямс, Джерри Крепс и Джефф Томпкинс, которые участвовали в её работе в различной степени. В октябре 1998 г. Джо Дассо и я приняли приглашение присоединиться к группе, и количество её членов возросло, таким образом, до 13.
ГИБ совместно с биолого-химическим факультетом в Свободном университете в Линчбурге (штат Вирджиния) 5-7 августа 1999 г. организовала и провела трёхдневную конференцию под названием «Бараминология-99: креационистская биология XXI века». На неё было приглашено 24 человека.
Официальные доклады были сделаны Тоддом К. Вудом, Д. Эшли Робинсоном, Куртом П. Визе, Питом Дж. Вильямсом и Полом Нельсоном. Среди тем, относящихся к бараминологии, были креационная биология, филогенетические заключения, библейские исследования, теория замысла, критерий гибридизации, эволюционные сети и несистематические поля. Последний вечер был посвящён семинару по бараминологии семейства, включающего в себя верблюдов. Эти доклады и дискуссии о верблюдах включали в себя введение, библейские и лингвистические исследования, данные палеонтологии, гибридизацию, молекулярные исследования, филогенетическое искажение и выводы. В последний вечер конференции состоялась организованная дискуссия на тему «Будущее ГИБ и бараминологии».
Была рассмотрена возможность учреждения нового общества и нового журнала. Участники добровольно взяли на себя ответственность за различные аспекты подготовки второй конференции на западном побережье в начале 2001 г.
Несмотря на некоторое расхождение во мнениях, у меня сложилось впечатление, что группа в целом была очень заинтересована достигнутыми на тот момент успехами и будущими перспективами бараминологии. Некоторые учёные предпочитали верить в древность Земли (миллиарды лет), но руководство и большинство присутствующих были едины в поддержке мнения о молодой Земле (тысячи лет). Введение в бараминологию и отчёт о конференции 1999 г. были написаны присутствовавшим ботаником Маргарет Хелдер, 1999.
Заключительные замечания
Бараминологию можно считать типологическим подходом к классификации форм жизни, и существующих ныне, и ископаемых. В прежние века учёные рассуждали типологически чаще, чем сейчас. Однако, из-за множества трудностей (например, конвергенций и отходов назад), которые не дают покоя мыслителю-макроэволюционисту, типологию принимают всё более широко.
Бараминологи считают, что они находятся на переднем крае современного прогрессивного мышления. Те, кому интересно узнать о десятилетней истории бараминологии больше, могут обратиться к источникам.
Благодарность
Дэвид Кавано, Курт Визе и Уолтер Ри-Майн прочитали ранние наброски этой рукописи и внесли много неоценимой информации. Помощь поступала также от Джона У. Куоццо, Нила Дорана, Скотта A. Мэхэти, Тодда Вуда и Д. Эшли Робинсона. В механических деталях помогал Лейн П. Лестер, управляющий редактор Ежеквартального журнала общества креационистских исследований (CRSQ). Статья была написана в ответ на неоднократные просьбы CRSQ. Редактор Эмметт Л. Вильямс.
Источники
CRSQ - Creation Research Science Quarterly
Bartz, Paul A. 1991. A refinement of biosystematics which reflects baraminic variation. CRSQ 28(1):18-20.
Cuozzo, Jack. 1998. Buried alive: the startling truth about Neanderthal Man. Master Books, PO Box 727, Green Forest, AR 72638.
___. 2000. Personal communications.
Denton, Michael. 1986. Evolution: a theory in crisis. Woodbine House, Rockville, MD.
de Queiroz, Kevin. 1992. Phylogenetic taxonomy. Annual Review of Ecology and Systematics 23:449-480.
___. 1997a. Misunderstandings about the phylogenetic approach to biological nomenclature: a reply to Liden and Oxelman. Zoologica Scripta 26(1):67-70.
___. 1997b. The Linnaean hierarchy and the evolutionization of taxonomy, with emphasis on the problem of nomenclature. Aliso (The Rancho Santa Ana Botanic Garden, Claremont, CA 91711-3157) 15(2):125-144.
Frair, Wayne. 1982. Serum electrophoresis and sea turtle classification. Comparative Biochemistry and Physiology 72B:1-4.
___. 1991. Original kinds and turtle phylogeny. CRSQ 28(1):21- 24.
___. 1999. Creationist classification--an update. Creation Matters 4(1):1,5.
Garcia-Pozuelo-Ramos, Celedonio. 1997. Taxonomy of primates. CRSQ 34(2):93-102.
___. 1998. Dental variability in the domestic dog (Canis familiaris): implications for the variability of primates. CRSQ 35(2):66-75.
___. 1999. Craniodental variability in the domestic dog (Canis familiaris) and its implications for the variability in primates. CRSQ 36(3):116-123.
Hartwig-Scherer, Sigrid. 1998. Apes or ancestors?: interpretations of the hominid fossil record within evolutionary and basic type biology. In William A. Dembski, editor. Mere creation: science, faith and intelligent design, pp. 212-235. Intervarsity Press, Downers Grove, IL.
Helder, Margaret. 1999. Introducing baramins. Creation Dialogue (5328 Calgary Trail South, Suite 1136, Edmonton, Alberta T6H 4J8) 26(3):6-7.
Hirayama, Ren. 1998. Oldest known sea turtle. Nature 392:705-708.
Iverson, John B. 1992. A revised checklist with distribution maps of the turtles of the world. Privately printed, Richmond, IN.
Klein, Richard G. 1999. The human career : human biological and cultural origins. The University of Chicago Press, Chicago, IL.
Lutz, Peter L., and John A. Musick. 1997. The biology of sea turtles. CRC Press, New York.
Marsh, Frank L. 1941. Fundamental biology. Published by the author, Lincoln, NE.
___. 1969. The form and structure of living things. CRSQ 6(1):13-25.
Milius, Susan. 1999. Should we junk Linnaeus? Science News 156(17):268-270.
O’Brien, Stephen J. 1997 (June). The human-cat connection. National Geographic 191(6):77-85.
ReMine, Walter J. 1990. Discontinuity systematics: a new methodology of biosystematics relevant to the creation model. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 207-216. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1993. The biotic message: evolution versus message theory. St. Paul Science, PO Box 28006, Saint Paul, MN 55128. [up]
___. 2000. Personal communications.
Robinson, D. Ashley. 1997. A mitochondrial DNA analysis of the testudine apobaramin. CRSQ 33(4):262-272.
Robinson, D. Ashley, and David P. Cavanaugh. 1998a. A quantitative approach to baraminology with examples from the catarrhine primates. CRSQ 34(4):196-208.
___. 1998b. Evidence for a holobaraminic origin of the cats. CRSQ 35(1):2-14.
Scherer, Siegfried. 1993. Typen des Lebens. Pascal Verlag, Berlin.
Williams, Pete J. 1997. What does min mean? Creation Ex Nihilo Technical Journal 11(3):344-352.
Wise, Kurt P. 1990. Baraminology: a young-earth creation biosystematic method. In R. E. Walsh, editor, Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345-360. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1992. Practical baraminology. Creation Ex Nihilo Technical Journal 6(2):122-137.
___. 1998. Is life singularly nested or not? In R. E. Walsh, editor, Proceedings of the Fourth International Conference on Creationism. Technical Symposium Sessions, pp. 619-631. Creation Science Fellowship, PO Box 99303, Pittsburgh, PA 15233-4303.
___. 1999-2000. Personal communications.
Можно приобрести распечатку
Печатный вариант статьи "Бараминология - классификация сотворённых организмов" (на английском языке) можно приобрести как отдельное издание . Цена $2.00 за экземпляр. Добавьте 15% за пересылку (минимум $3.00, максимум $10). Отправьте заказ по адресу: Creation Research Society, P.O. Box 8263, St. Joseph, MO 64508-8263.
[Если эта информация была вам полезна, просим Вас с молитвой рассмотреть возможность пожертвования для того, чтобы сделать это служение по наставлению в вере доступным для Вас и Вашей семьи! Пожертвования снижают налоги].
Автор: доктор философских наук Уэйн Фрэр. Текст опубликован в Creation Research Society Quarterly, Vol. 37, No. 2 (September 2000), pp. 82-91. Предоставлено Обществом креационистских исследований.Переводчик : Yevgheny Yeryomin
Copyright © 2001, Creation Research Society, Все права защищены – кроме того, что помечено на приложенной странице "Пользование и авторские права", которая даёт пользователям ChristianAnswers.Net обширные права на использование этой страницы у себя дома, в личном свидетельстве, в церквах и школах. Все фотографии предоставлены Films for Christ и защищены авторским правом.
Узнайте больше о Сотворении в нашей СуперБиблиотеке
 Индекс
ИндексДОМАШНЯЯ СТРАНИЦА и УКАЗАТЕЛЬ
ChristianAnswers.Net
Christian Answers Network
PO Box 577
Frankfort KY 40602 USA
Присылайте свои вопросы